
综述丨ISME J(IF:10.8): 细菌-真菌竞争土壤资源的机制及意义(国人佳作)
- 看不见的线
- 11282
- 2024-07-22 09:01:47
- 文章来源:微生态
导读
阐明细菌和真菌之间的复杂相互作用,这些相互作用决定了土壤中微生物群落的结构、组成和功能,并调节碳(C)和养分通量,对于理解生物地球化学循环至关重要。在各种相互作用中,资源竞争是决定土壤中这两大微生物类群适应和生态位分化的主要因素。这是因为微生物生长的碳和能量限制是一种规则,而不是例外。本文综述了细菌和真菌对碳、能量的需求,它们对这些资源和其他资源的竞争机制,导致生态位分化,以及全球变化对这种竞争的影响。标准化微生物利用偏好表明,细菌利用简单有机化合物的效率比真菌高1.4-5倍,而真菌在利用复杂化合物方面的效率要比细菌高1.1-4.1倍。因此,细菌在利用简单底物方面比真菌更胜一筹,而真菌则利用复杂化合物。细菌还与真菌竞争复杂底物降解过程中释放的产物。在此基础上,对这两个类群在土壤中的空间、时间和化学生态位进行了划分。在CO2浓度升高、氮沉降、土壤酸化、全球变暖和干旱等五大全球变化的影响下,竞争将会加剧。CO2浓度升高、氮沉降和全球变暖增加了细菌的优势,而土壤酸化和干旱增加了真菌的竞争力。
论文ID
原名:Mechanisms and implications of bacterial-fungal competition for soil resources
译名:细菌-真菌竞争土壤资源的机制及意义
期刊:The ISME Journal
IF:10.8
发表时间:2024.5
通讯作者:Chaoqun Wang,Yakov Kuzyakov
通讯作者单位:山东农业大学农学院;德国哥廷根大学
DOI号:10.1093/ismejo/wrae073
综述目录
1 引言
2 细菌和真菌的碳、能量需求
3 细菌-真菌竞争与生态位分化
3.1 细菌-真菌竞争机制
3.2 生态位分化
4全球变化影响
4.1 CO2升高
4.2 氮沉降
4.3 酸化
4.4 全球气候变暖
4.5 干旱
4.6 其他全球变化因素
5 细菌和真菌共存
6 结论与未来展望
主要内容
1 引言
就生物多样性、生物量及其对生物地球化学过程的影响而言,细菌和真菌是土壤中的关键生物成分。它们在土壤中总是共存,并形成复杂的相互作用,这对它们的生存、适应、建立、维持和功能至关重要。在经典生态学理论中,这些普遍存在的相互作用可分为互利共栖、竞争、共生、寄生、偏害共栖和无关共栖。在这些互作类型中,资源竞争主导了土壤中的这些相互作用。因此,微生物适应主要涉及竞争成功。这是因为对有限空间和资源(例如,碳(C)和能源、营养物质、水)的竞争无处不在。竞争也是增加微生物群落稳定性的重要机制,通过容纳各种代谢冗余物种和限制微生物病原体的过度生长。除了对微生物群落结构的影响外,细菌-真菌竞争还关键地调节了多种生态系统功能,如养分循环;凋落物、根际沉积物和土壤有机质(SOM)的分解;土壤结构形成、提高土壤肥力;抑制植物病害、支持植物生产力以及增强生态系统的抵抗力和恢复力。尽管人们越来越意识到细菌和真菌具有影响全球生物地球化学和多种生态系统功能的巨大能力,但它们之间的竞争在土壤微生物组研究中常常被忽视。
由于细菌和真菌对有机化合物和营养物质以及对相同土壤位置(生境)的需求有很大的重叠,因此对碳和能量的竞争比对其他资源的竞争更强。重要的是,对能量的竞争(主要储存在植物光同化的有机化合物中)可能远远强于对其他资源的竞争。这与土壤中微生物碳利用效率(CUE)通常<0.4这一事实相符合,表明>60%的碳用于通过将有机化合物氧化为CO2来获得能量,而不是用于结构碳。此外,微生物利用相当多的碳来合成储能物质,许多微生物过程需要能量而没有碳投入,导致微生物能量利用效率(EUE)低于CUE。在竞争相互作用中,两个群体必须始终分配能量以获得最有限的资源,并克服竞争的负面影响。这使得理解控制细菌-真菌对碳和能源竞争的机制对于解开碳循环和稳定的生物地球化学过程至关重要。
微生物竞争包括两种主要类型:剥削性竞争(也被称为争夺性竞争),其中一个种群迅速消耗另一个种群所需的有限资源,而两个种群之间没有直接的相互作用;干扰性竞争(又称竞赛比赛),竞争种群产生直接的拮抗相互作用,其中一个种群通过竞争成功占有资源。细菌和真菌形成了相互利用和干扰竞争的各种机制,调节这两大群落之间的生态位分化。细菌通常以快速吸收不稳定的碳和能量资源为特征,而真菌有效地利用更难降解的碳源。真菌可以通过形成菌丝和菌丝体来扩大其空间生态位,以获取C和营养资源。
各种生物因素(例如,植物种类、根-微生物相互作用、微生物多样性和密度等)和非生物限制因素(如C、能量、养分的有效性、pH、湿度、通气性、温度等)可以影响土壤中的竞争。各种全球变化过程(例如,CO2浓度升高、氮沉降、土壤酸化、气候变暖、干旱等)强烈影响这些生物和非生物因素,从而调节竞争。例如,在CO2浓度升高的情况下,植物生长和碳输入(例如,根际沉积物和根系分泌物)通常会增加,这可能会降低细菌-真菌对C和能量的竞争强度。由于细菌和真菌对营养物质的需求(如C:N:P化学计量)以及对温度、pH、湿度和氧气浓度的敏感性不同,它们对全球变化的响应具有界特异性。因此,即使生物地球化学环境发生轻微变化,也可能对微生物代谢以及对C和能源的需求产生强烈影响。因此,全球变化强烈地影响着竞争力。考虑到生态系统的稳定性和功能在很大程度上取决于细菌和真菌的性能和平衡,理解和预测这种竞争对全球变化的响应是最紧迫的研究问题之一。我们迫切需要确定的是,在变化的气候条件下,细菌-真菌对C和能量的竞争及其对生物地球化学过程的影响如何变化。反过来,这将有利于保护、管理和减轻生态系统的抵抗力和恢复力。
由于气候变化,全球范围内SOC分解的异养呼吸增加,从而增加了大气中CO2的输入。然而,由于植物生长和土壤微生物自养固定作用的增加,土壤C输入的增加可以抵消土壤C向大气中的流失。此外,SOC分解的温度敏感性取决于植物凋落物的数量和化学性质以及先前存在的SOC。因此,即使在特定的生物群落中,当地的生物地球化学环境也会强烈影响微生物代谢对气候的响应。
本篇综述评估了细菌和真菌对土壤中碳源的需求和偏好。然后,总结了竞争机制以及由此导致的生态位分化。最后,概述了在真实土壤条件下,各种全球变化对细菌-真菌竞争碳和能源的影响。
2 细菌和真菌的碳、能量需求
2.1 细菌和真菌的碳、能量来源
细菌和真菌生长、维持和休眠期间的所有过程都消耗能量,其中大部分消耗C。每个群体对有机化合物的偏好取决于它们的结构复杂性和表面性质,部分取决于代谢特性。细菌和真菌都可以利用土壤中的各种有机化合物来获取C和能量,以维持自身的生长和维持。根据来源,主要的C和能量来源可分为根际沉积物、植物凋落物、微生物残体、有机肥和SOM(表1)。
据估计,植物C向根际沉积物的分配为光同化碳的10%-50%,相当于多年生和一年生植物的800~4500 kg Cha-1year-1。重要的是,根际沉积物包括根系分泌物:各种可溶性低分子量化合物,特别是糖类、羧酸和氨基酸,它们是重要的碳和能量来源,因为它们是可溶的,因此非常容易被细菌和真菌利用,因此,它们几乎不需要额外的溶解和吸收成本。此外,这些化合物中的许多是代谢循环中的关键物质。
植物凋落物的微生物利用取决于分解阶段。在早期阶段,通过各种过程从细胞中浸出的可溶性和不稳定分子(如糖、氨基酸等)会被细菌和真菌迅速吸收。在凋落物分解后期,剩余的复杂和难降解的化合物(如木质素、单宁和几丁质)需要较高的能量和C输入,以产生水解和氧化所必需的胞外酶。微生物残体C占表层土壤有机碳总量的35-51%,是潜在的重要碳源和能源。微生物残体的解聚和分解可能比植物残体中复杂化合物(如木质素)的分解更快,因为有机化合物更小,具有更高的C:N比(蛋白质和氨基糖占优势)。
土壤中主要C和能量来源的NOSC值按以下顺序增加:脂类、微生物残体、木质素、氨基酸、酚类、植物凋落物、NaOH可提取SOM、糖类、HCl可提取SOM和羧酸(图1)。尽管有机化合物中的能量含量随着NOSC值的增加而减少,但能量可用性增加。因此,微生物优先利用NOSC值较高的有机化合物。
表1 土壤中细菌和真菌的碳、能量来源。
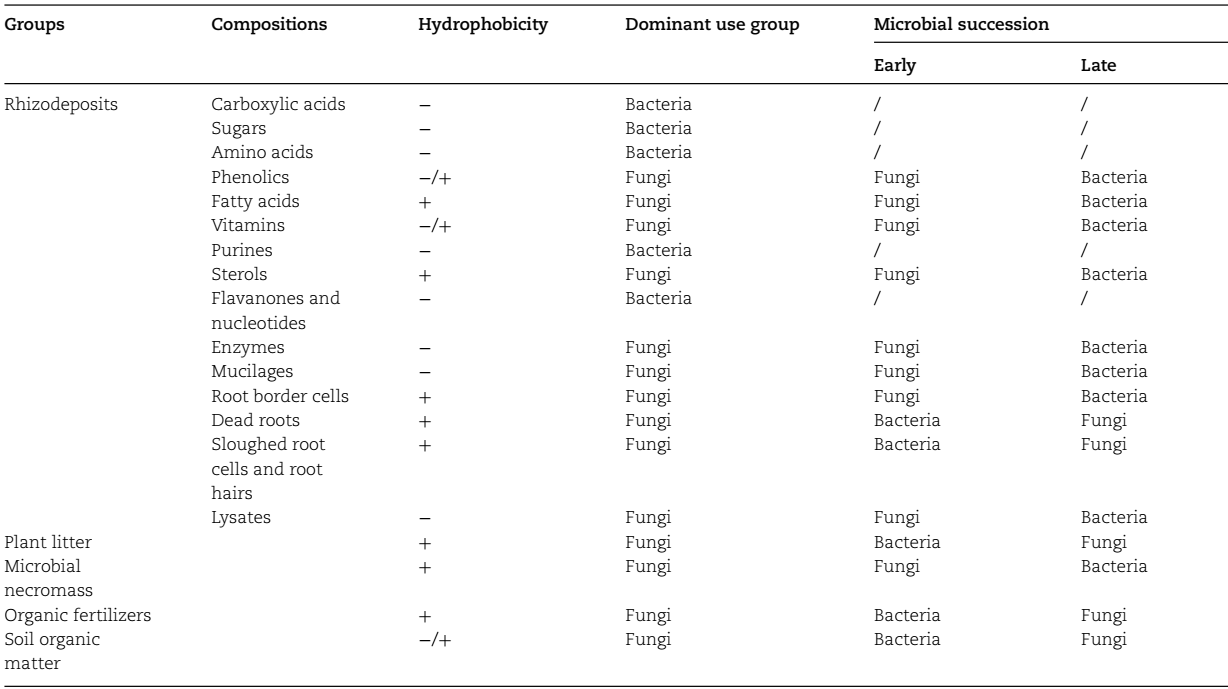

图1 作为土壤中微生物的碳和能量来源的主要化合物中C原子的名义氧化态(NOSC) (X轴)和能量含量(Y轴)的概念表示。
2.2 碳源和能源的疏水性
一般来说,细菌会迅速吸收不稳定的有机化合物,而真菌通常与难降解的形式有关。除了许多简单的化合物(葡萄糖、甘氨酸、乙酸)外,一些复杂的化合物(淀粉)主要被细菌吸收。同样,在桦树-柳树系统中,真菌从N-乙酰葡糖胺利用中吸收的C是细菌的2倍,真菌 :细菌 PLFA比率仅为0.14。
微生物对C的利用不仅取决于其结构的复杂性和能量的可利用性,还取决于有机化合物的溶解性。这是因为溶解性决定了有机化合物对微生物的可及性。可溶性有机化合物通过溶液扩散,使微生物能够从较大的土壤体积中捕获它们。相比之下,不溶性化合物通常是高分子量化合物,或吸附在金属氧化物、粘土矿物或有机物上,使其不易被微生物摄取。与不溶性化合物相比,可溶性化合物更有效地被胞外酶水解或氧化,导致细菌和真菌之间的激烈竞争,尤其是可溶性化合物。微生物必须分配相当大的能量来分解不溶性有机化合物和土壤矿物之间复杂的化学(如配体交换)和/或物理(如静电引力、疏水分配)联系。这极大地降低了以不溶性有机化合物作为底物的微生物CUEs和EUEs。真菌比细菌具有更高的能力将能量从有机化合物转化为其生物量,这可能会增加它们利用不溶性化合物的竞争力。
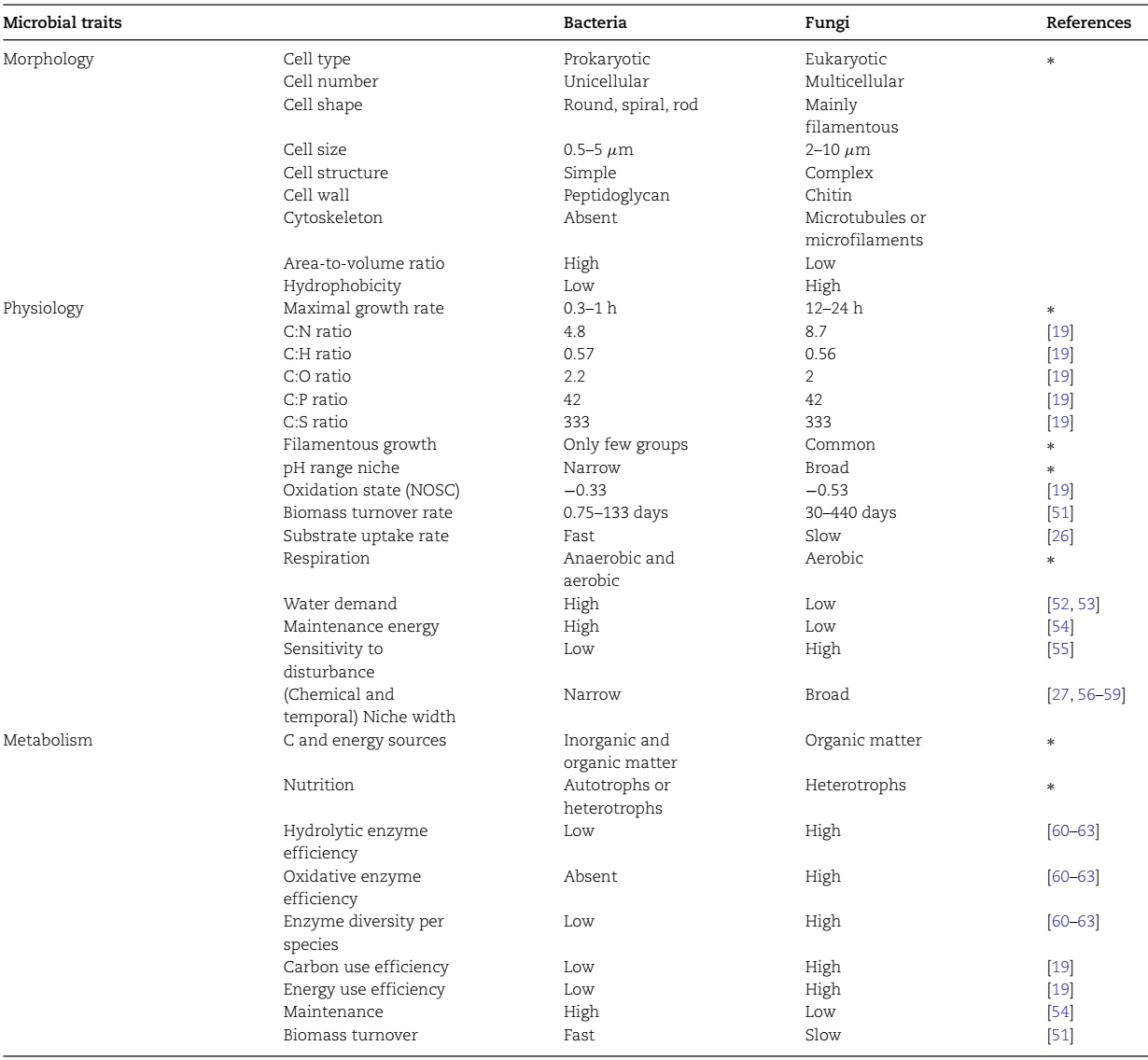
基于结构复杂性和表面疏水性,有机化合物可以分为亲水性化合物和疏水性化合物。具有疏水基团和官能团(例如,-OCH3、-CH3、-CN)的有机化合物从土壤溶液中到矿物上的疏水分配抵消了微生物的分解作用。有机化合物的疏水性越高,其微生物分解就越慢。因此,细菌和真菌都优先吸收和利用亲水性有机化合物作为其碳和能量来源。细菌的形态特征(例如,简单的细胞结构、大的面积/体积比)和生理特征(例如,快速生长和底物吸收速率)有助于它们快速利用亲水性化合物(表2)。真菌比细菌具有更高的疏水性和更有效的胞外酶(表2),使它们能够通过氧化和水解来利用疏水性化合物。
表2 土壤中细菌和真菌适应的生理和代谢特性的主要差异。
2.3 对碳源和能源的偏好
为了评估对碳源和能源的偏好,收集了155个关于将13C标记物掺入微生物生物标志物-磷脂脂肪酸(PLFAs)的数据。将真菌相对于细菌PLFAs的13C富集(13CFungi/13CBacteria)标准化为真菌相对于细菌的生物量C含量。较高的MUP(微生物利用偏好)表明真菌对底物C的同化作用高于细菌。当MUP值大于1时,真菌对底物的竞争大于细菌,反之亦然。
MUP值为0.20-0.72对应于细菌对小分子(MW<200 Da)和亲水性化合物的竞争力比真菌强1.4-5倍(图2A)。相比之下,复杂底物(例如,植物残体、微生物残体、蛋白质、纤维素、纤维二糖、生物炭等)的MUP值在1.1-4.1之间(图2B)。因此,细菌在简单底物方面胜过真菌,而真菌在复杂底物方面具有主要优势。
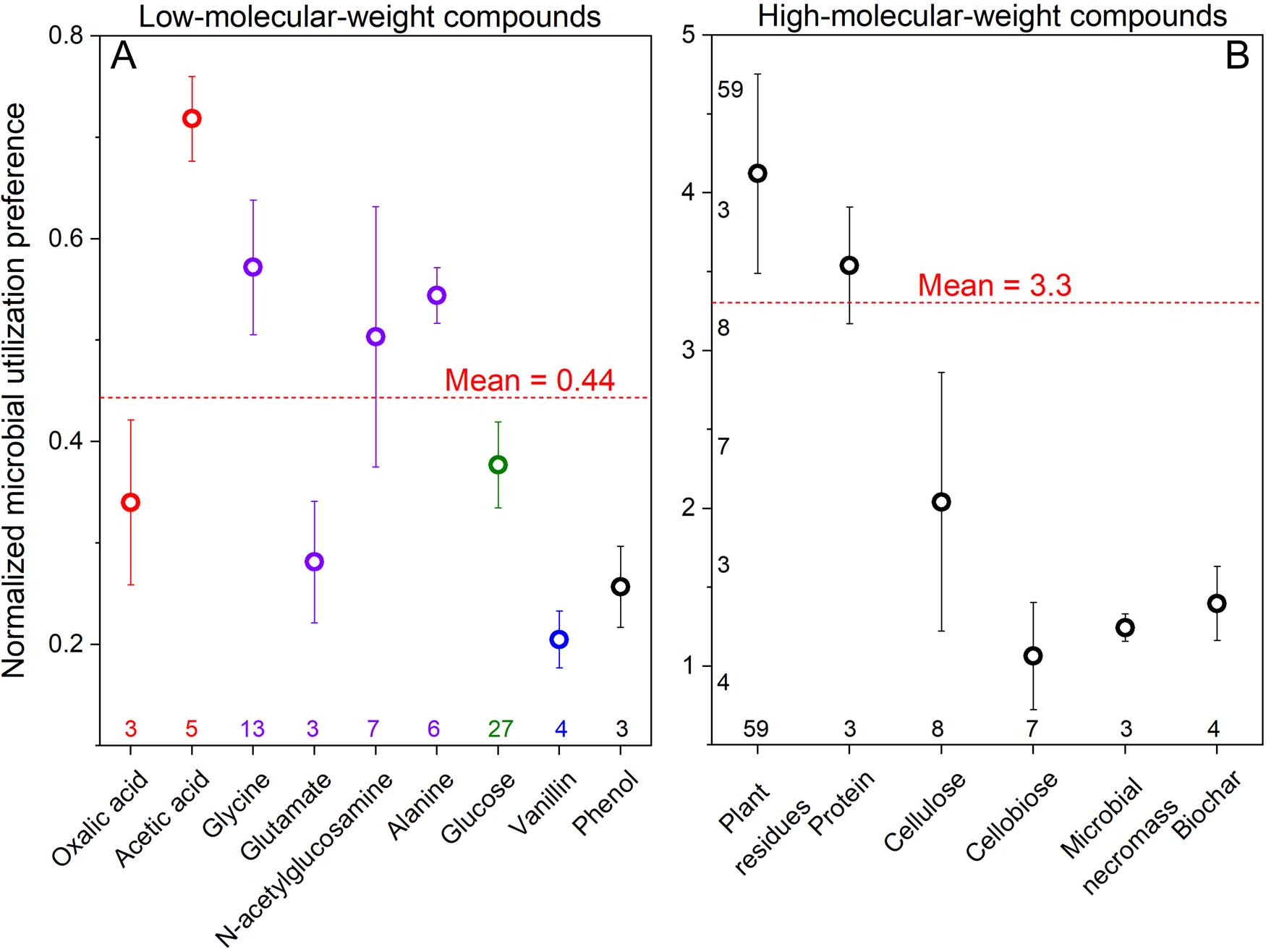
图2 以各种简单(A)和复杂(B)化合物作为底物,掺入真菌PLFAs相对于细菌PLFAs的标准化微生物利用偏好(MUP)。红色虚线代表利用简单(A)和复杂(B)化合物作为底物的平均MUP值。MUP值越接近1,细菌和真菌对底物的竞争力越相似。
随着分解时间的延长,以植物残体作为底物的MUP逐渐降低(图3),表明真菌是此类残体更好的竞争者,尤其是在分解早期阶段。然而,后来,细菌直接或间接地从真菌中获得更多的产物,反映了交叉喂养的普遍现象(图4)。这是因为细菌只能利用植物残体分解产生的较小化合物,需要时间才能发生。这也得到了纤维素和纤维二糖作为底物(具有较快的降解速率)的MUP低于使用慢降解蛋白质和植物残体的MUP的支持(图3)。
交叉喂养通常假设一个生物的代谢产物(有机化合物和营养物质)被另一个生物利用,因此通常只考虑细胞内代谢。然而,土壤中的交叉喂养包括并可能主要涉及一个微生物群体利用另一个群体产生的胞外酶的细胞外反应释放的产物。考虑到胞外酶的反应产物通过扩散作用的全方位损失,我们假设土壤中交叉喂养涉及机制2:底物交叉喂养(图4)。虽然交叉喂养相互作用很可能是正面的,但它们在生物体之间产生了竞争。底物交叉喂养是生物体对产物进行剥削性竞争的基础。虽然细菌和真菌在其他交叉喂养相互作用中不竞争产物(图4),但它们在一定程度上竞争其他共享资源(如水、氧气、营养物质等)。
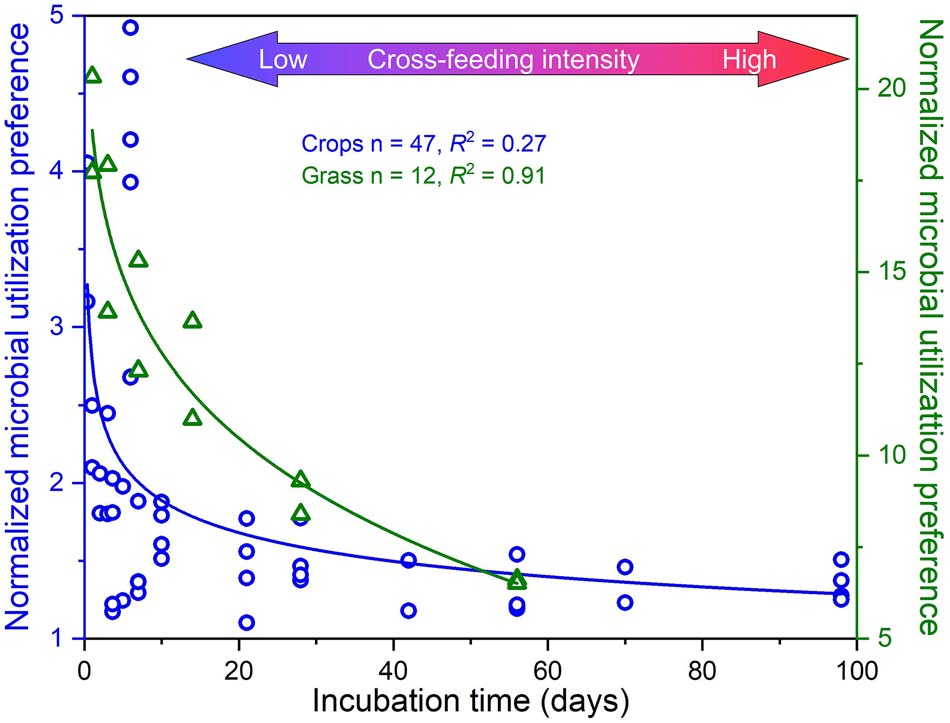
图3 以作物残体(蓝色圆圈)和草残体(绿色三角形)为底物,将C掺入真菌PLFAs相对于细菌PLFAs的标准化MUP与培养时间(即残体分解时间)的关系。MUP值随残体分解而急剧下降,表明细菌和真菌之间存在交叉喂养相互作用。
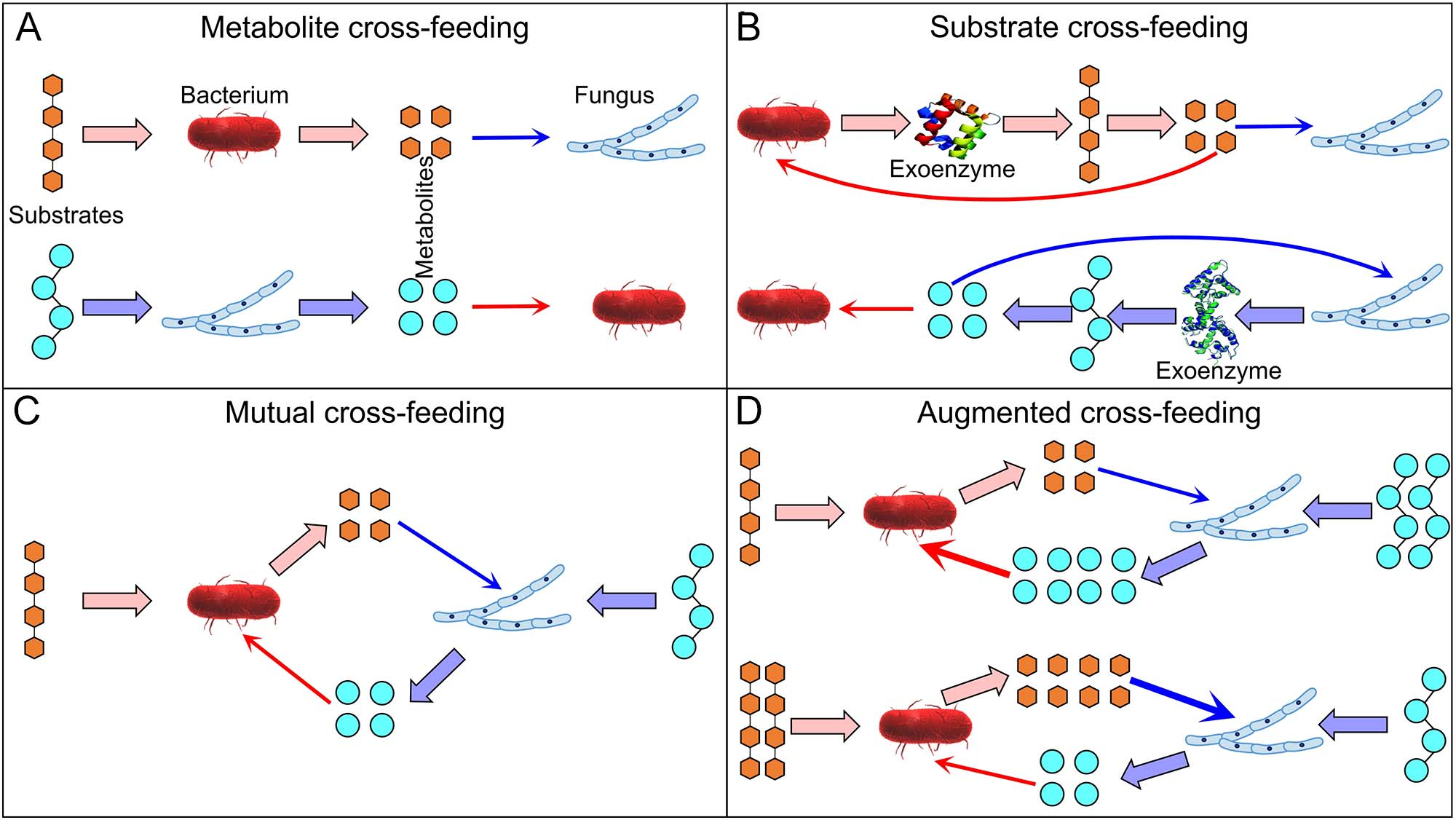
图4 细菌-真菌交叉喂养类型。
3 细菌-真菌竞争与生态位分化
土壤中有限的碳和能量可利用性是规则而非例外,因此通常会限制所有非丝状细菌、放线菌和真菌的生长。土壤中的这些限制导致细菌不断与真菌竞争有机碳和能量。对能量的竞争比对碳本身的竞争要激烈得多。首先,微生物可以在胞内和胞外循环C,这需要能量的投入,而不是新的C。其次,微生物必须投入能量来减少有机和无机化合物,这发生在没有C利用的情况下。第三,微生物分配大量的能量用于维持,各种过程(例如,细胞分裂、代谢变化、细胞运动、基因表达调控、能量溢出反应等)消耗能量,但不消耗或很少消耗C。例如,在30℃的有氧条件下,细菌的维持能量(12 kJ mol-1C h-1)比真菌(8 kJ mol-1C h-1)高50%。然而,这两个群体都需要更多的能量来相互竞争,从而减少了用于其他功能的能量;这反过来又增加了能源竞争的强度。
细菌和真菌强烈竞争易获得的碳和能源,但它们也进化出了分解难降解有机化合物的竞争和互惠策略。革兰氏阴性菌通常是快速生长的r-策略菌,可以快速吸收容易获得的底物,而真菌和革兰氏阳性菌是缓慢生长的K-策略菌,可以有效地利用难降解的有机化合物。这反映在根际的细菌数量(尤其是革兰氏阴性菌株),比根际的真菌数量高得多,而在根际中,根系不断地分泌简单的底物。当向土壤中添加葡萄糖时,革兰氏阴性菌的丰度增加,而真菌和革兰氏阳性菌的丰度随着难降解有机物的添加而增加。真菌/细菌比值在添加易获得的碳源(如糖、氨基酸)后总是降低,但随着难降解有机物的增加而增加。
这两个群体共存于土壤中的多个生态位(表3),增加了它们对C和能量的竞争。生态位分化有效地减少了这种竞争。生态位分化的胜利者取决于它们的形态(例如,细胞大小、结构、疏水性等)、生理(例如,生长速度、水分和养分需求)和代谢(如酶催化效率,C和能量利用效率)特性(表2)。生态位的理化条件(如有机化合物的复杂性、O2浓度、pH等)也起到一定的作用。
表3 细菌与真菌的生态位分化。
3.1 细菌-真菌竞争机制
存在两类竞争类型:剥削性竞争和干扰性竞争。细菌和真菌已经发展出了各种策略,通过两种类型和生态位分配来相互竞争土壤中的碳和能量资源(图5)。
在剥削性竞争中,细菌在容易获得的有机化合物的竞争中胜出(图5)。与真菌细胞相比,细菌细胞体积较小,其表面积/体积比更大,从而使细菌能够更快地接触和吸收小分子溶解性有机化合物(表2)。细菌的生长速度要快得多(表2),使其能够更迅速地占据有机物资源。最后,细菌的细胞膜结构比真菌简单,有利于直接从土壤溶液中摄取小分子化合物。
相比之下,真菌对复杂有机化合物的竞争更胜一筹(图5),主要是因为它们能高效地产生几乎所有的胞外酶来分解这些结构。真菌可以利用比细菌更丰富的碳和能源,因为它们的菌丝增加了栖息地和探索体积(表2),而单细胞细菌仅限于土壤中的小体积。菌丝体从遥远地点转移物质和缓慢生长(表2)有助于真菌在土壤中保持更稳定的状态,提供比细菌更多的时间来分解复杂化合物。
在干扰性竞争中,两个群体都产生化合物来抑制或杀死对方(图5)。细菌可以产生细菌素(一组小的抗菌肽或蛋白质)和抗真菌化合物,减少甚至阻止真菌生长。反过来,真菌可以产生抗生素(如青霉素、链霉素、四环素等)和真菌毒素来限制细菌生长。两个群体产生的挥发性化合物通过充满空气的孔隙扩散,并抑制竞争者的活性和生长。
两个群体都可以形成生物膜,通过排除竞争对手来竞争当地的碳和能源(图5)。由于化合物向外扩散缓慢,生物膜通过积累拮抗分子来增加竞争力。生物膜还可以保护封闭的细菌或真菌免受捕食者的侵害,并促进某些物种向富含C的区域生长。群体感应是许多物种通过调节胞外聚合物的产生、表面附着、运动和扩散来形成生物膜的常用策略。通过群体感应,微生物种群可以共同应对竞争,例如通过增加抗菌化合物的产生和改变它们的行为来获得竞争优势。
细菌和真菌都会产生特定水解酶(例如,几丁质酶、蛋白酶等)来破坏其竞争者的细胞壁或蛋白质。这两个群体都会产生铁载体,以螯合土壤中的铁,从而使其竞争者挨饿,而这些竞争者需要铁才能生长和生存。
微生物可以通过改变环境的理化性质来间接地相互抑制(图5)。例如,在Collimonas fungivorans和黑曲霉(Aspergillus niger)之间的竞争性相互作用中,前者产生酸,为后者创造了不利条件。一些微生物可以充当其他微生物群的捕食者或寄生虫,导致直接的生存竞争。蘑菇食用使某些细菌能够捕食活的真菌菌丝以获得营养物质和C。例如,细菌属Collimonas在侵入土壤中的Absidia菌丝后,其数量在两周内增加了4倍。
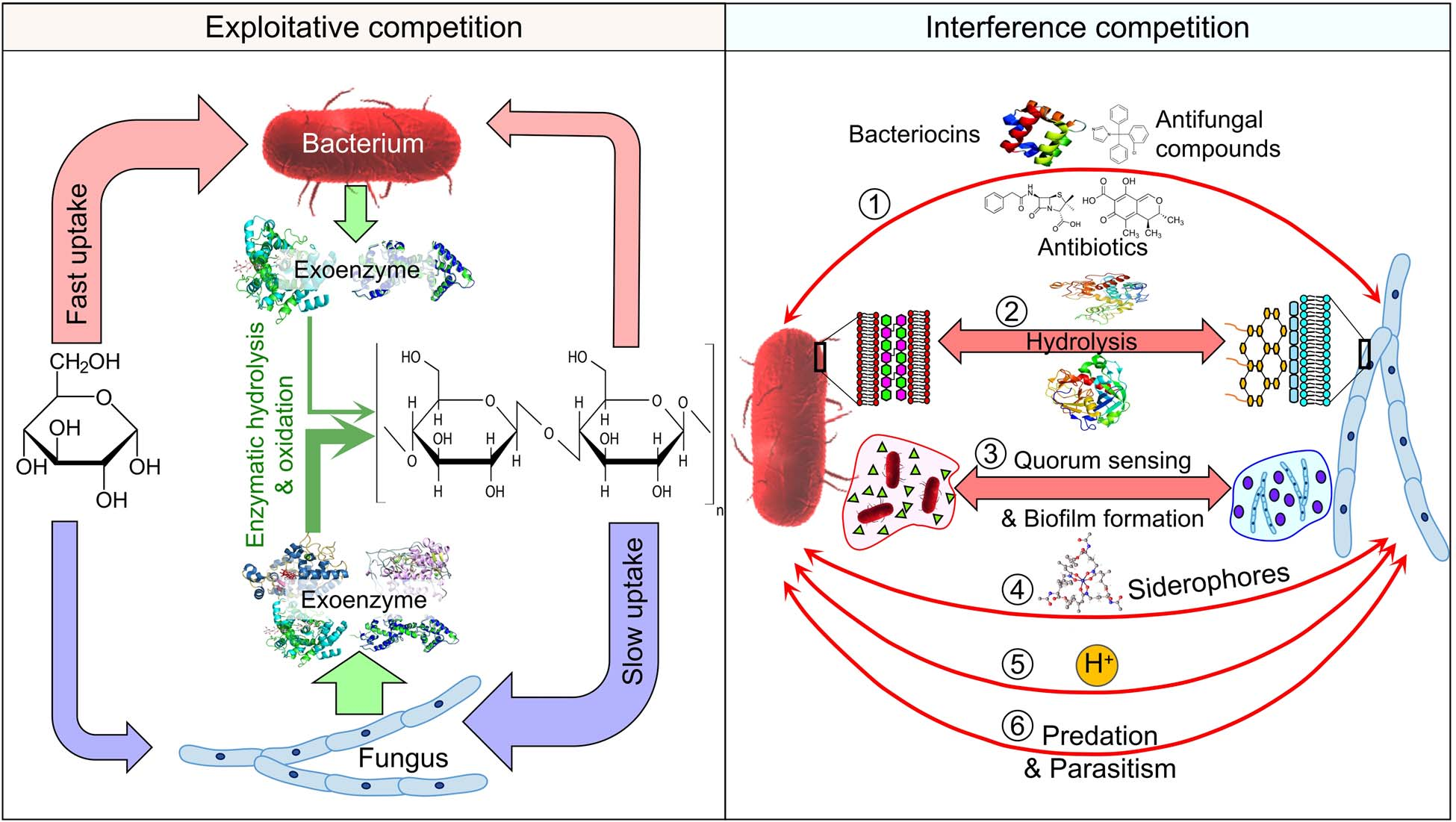
图5 细菌和真菌在剥削性(左)和干扰性(右)竞争中争夺碳源和能源的策略。在剥削性竞争中,细菌快速吸收易获得的化合物,从而胜过真菌,而真菌则依靠其具有高催化效率的多种酶在复杂化合物方面胜过细菌。在自上而下的干扰性竞争中采取的相互竞争机制包括:(i)产生各种化合物来抑制或杀死对方;(ii)分泌特异性水解酶(例如,几丁质酶、蛋白酶等)来破坏竞争者的细胞壁或蛋白质;(iii)产生铁载体以从土壤中螯合铁(和其他多价阳离子),从而使那些需要铁才能生长和生存的竞争者挨饿;(iv)形成生物膜以提高竞争力;(v)释放H+离子,使土壤酸化,为竞争者创造不利条件;(vi)直接捕食或寄生竞争者。
3.2 生态位分化
生态位分化是竞争相互作用减少的结果和机制。这里区分了化学、空间和时间生态位分化(图6)。
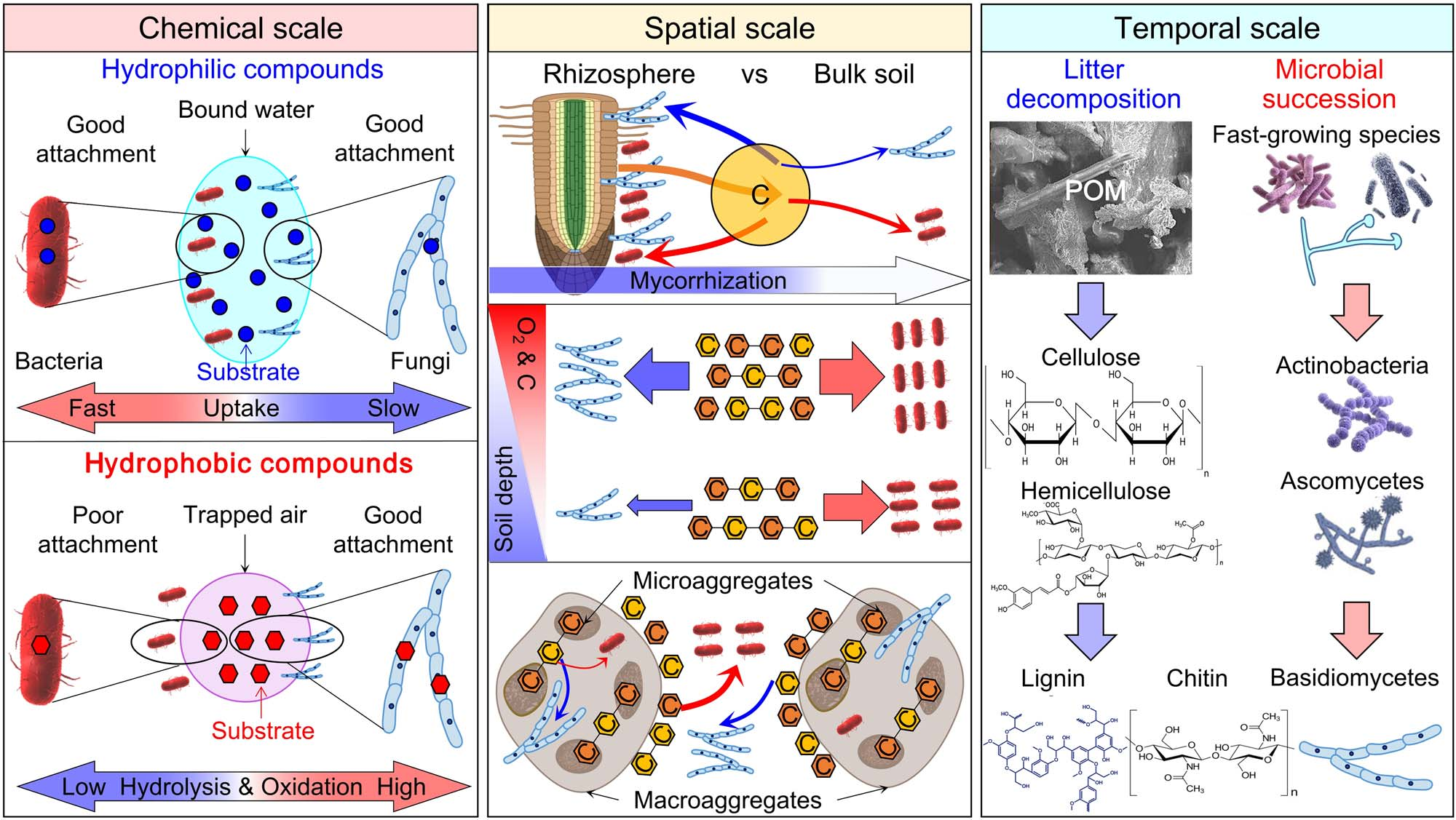
图6 细菌和真菌在化学(左)、空间(中)和时间(右)尺度上的生态位分化。在化学尺度上,细菌能够迅速摄取亲水性有机化合物,从而胜过真菌。真菌由于其更有效的胞外酶和菌丝在疏水表面上更好的附着在竞争疏水性有机化合物方面优于细菌。在空间尺度上,(i)真菌在根际比在非根际土壤中更具竞争力,因为根际沉积降低了竞争强度和菌根真菌在根系的定殖;(ii)细菌对碳的竞争随着土壤深度的增加而超过真菌;(iii)细菌在大团聚体中比真菌更有竞争力,而真菌在微团聚体中比在大团聚体中更有竞争力。在时间尺度上,植物凋落物分解过程中的微生物演替遵循如下顺序:快速生长的微生物,其次是具有纤维素分解、半纤维素分解和果胶分解能力的真菌和细菌,最后是能够分解木质素和几丁质的真菌。
3.2.1 化学生态位分化
在有机化学方面,细菌在利用亲水性化合物方面胜过真菌,而真菌则更适合利用疏水性化合物(图6)。在高地和水稻土壤中,与13C标记的玉米秸秆一起培养2天后,亲水性化合物在细菌磷脂中的掺入量是真菌磷脂的16倍。相比之下,疏水性化合物在真菌磷脂中的掺入量是细菌磷脂的1.5倍。
细菌对亲水性化合物的竞争主要是因为它们的快速生长和较大的表面积/体积比(表2),这有利于在富含亲水性化合物的土壤表面定殖。具有较大NOSC值的有机化合物亲水性更强(即更多的-COOH和=C=O基团),因此易于被微生物从土壤溶液中吸收。重要的是,亲水性化合物具有较高的能量可用性,因为它们已经是可溶性的,并且它们通过细胞膜被微生物吸收所需要的能量低于疏水性化合物。
从元素化学计量的角度来看,由于细菌生物量中的C:N比比真菌生物量中的C:N比低2倍,细菌必须在亲水性富氮化合物竞争中胜过真菌。通常,亲水性化合物(例如,氨基酸、多肽、氨基糖)的C:N比低于疏水性化合物。细菌在植物源N的短期同化中占优势也印证了这一点。
真菌依靠其广泛而有效的胞外酶(表4)和菌丝在疏水表面的更好附着来竞争疏水性化合物(图6)。复杂的疏水性化合物必须被胞外酶分解和/或氧化后才能被摄取。大多数疏水性化合物解聚的第一步是由胞外氧化酶(例如,过氧化物酶、酚氧化酶、漆酶等)催化氧化。疏水性化合物的氧化产生更多的-COOH、=C=O和≡C-OH基团,增强了它们的亲水性。虽然有些细菌可以产生氧化酶,但其酶活性和丰度远低于真菌(表4)。因此,真菌主导疏水性化合物的分解。
微生物必须投入大量资源(能量、C、N、P、S),才能使植物凋落物和微生物残体中的疏水性化合物(即许多-CH3基团、脂肪族、芳香族)得到利用。这就需要在合成和释放胞外酶方面投入相当大的能量。例如,在150 d的植被周期内,合成β-葡萄糖苷酶的能量成本为1.9-14×10 -6 J g-1soil,假设β-葡萄糖苷酶含量为2 ng g-1soil,每1 mol氨基酸聚合需要7.1 mol ATP。在真实的土壤条件下,微生物比β-葡萄糖苷酶多投入至少两个数量级的能量来产生所有参与分解聚合物的酶。因此,胞外酶的产生降低了从有机化合物中获得的净能量。然而,胞外酶只有在其底物浓度较高和/或胞外酶能有效地将底物中储存的能量转化为微生物生物量以抵消其生产的能源成本时才是有益的。具有较低NOSC值的疏水性化合物具有较高的Gibbs自由能(图1),真菌使用各种底物的EUE均高于细菌(表2)。当竞争疏水性化合物时,这是一个优势。
微生物细胞需要与疏水性化合物接触才能利用它们。真菌分泌疏水蛋白,随后形成两亲性蛋白膜,以促进气生菌丝的形成和运动。疏水蛋白膜可以使菌丝附着在有机化合物的疏水表面(图6)。一些真菌(如Candida、Aspergillus、Ustilago、Trichosporon)可以高效产生表面活性剂,以降低其细胞表面张力,削弱细菌细胞在疏水表面的附着,这些策略有助于真菌在疏水性有机化合物竞争中胜过细菌。
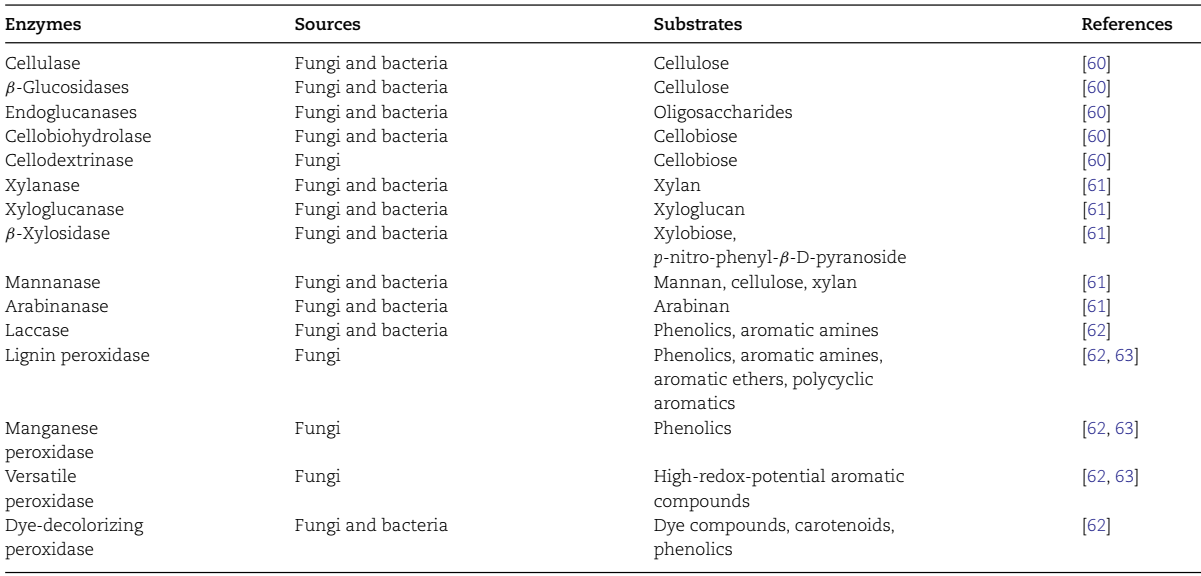
表4 主要纤维素降解酶和木质素降解酶的特性。
3.2.2 空间生态位分化
有机化合物的空间异质性(如根际vs.非根际土壤、表土vs.底土、大团聚体vs.微团聚体)(表3)导致空间生态位分化(图6)。在根际和非根际土壤中,细菌对容易获得的碳和能源的竞争力均高于真菌,但真菌在根际的竞争力高于非根际土壤(图6)。这被前者的真菌/细菌比率高于后者所证实。这是因为各种菌根类型占据根表面,并发挥着至关重要的作用,特别是在根际,捕获根系释放到土壤中的分泌物。
相对于非根际土壤,根际主要选择富营养细菌。根际富营养细菌比例高达7倍证实了这一点。根际中这些细菌的高丰度同时加速了碳的消耗,并降低了寡营养细菌对碳的竞争力。因此,根际真菌的相对竞争力高于非根际土壤,因为它们直接从根系获得能量(图6)。相比之下,真菌依靠其高效的胞外酶系统,在富含复杂化合物的生态位中胜过细菌(表3)。
新鲜植物来源的C和SOM的数量随土壤深度的增加而减少,导致细菌和真菌之间的生态位分化(图6)。SOM的能量含量随深度的增加而强烈降低,例如,在0~20 cm和20~100 cm土层,分别为630~1800 GJ ha-1和280~4100 GJ ha -1。然而,由于难降解但富含能量的疏水性化合物(木质素衍生物、脂肪酸、脂类等)随深度的增加而积累,单位C的能量含量随土壤深度的增加而增加。相反,革兰氏阳性菌的比例随着土壤深度的增加而增加,而革兰氏阴性菌和真菌的比例则降低,从而降低了真菌/细菌比值。这是因为深层土壤中孔隙较小的压实土壤结构部分被水填充,降低了O2浓度,从而减缓或阻止真菌(好氧生物)生长。同样,在缺氧生态位中,细菌比真菌占优势(表3)。深层土壤中较少或没有根系会大大减少植物与真菌之间的寄生或共生。因此,这些机制会随着土壤深度的增加而降低真菌对有机化合物的竞争力(图6; 表3)。
不同粒级团聚体中C和能量来源的异质性分布也导致细菌和真菌之间的生态位分化(图6)。易利用有机化合物通常在大团聚体中比在微团聚体中更丰富,而难降解有机化合物则受到微团聚体的强烈保护(图6)。微团聚体中有机物与矿物的强联结性限制了微生物的可及性。因此,大团聚体中C和能量来源的竞争更强(图6)。在这种较大的团聚体中,细菌和真菌之间的负连接数量高达2倍。与微团聚体相比,大团聚体中高达3.6倍的正细菌-细菌相互作用使细菌比真菌更有竞争力。因此,快速生长的细菌在大团聚体中的微生物群落中占主导地位(表3)。大团聚体中变形菌门和拟杆菌门(主要利用活性碳)的丰度高于微团聚体,酸杆菌门、绿弯菌门和疣微菌门(具有寡营养生活策略)的丰度低于微团聚体。
3.2.3 时间生态位分化
在植物凋落物分解早期细菌的优势和后期阶段真菌的优势有力地支持了时间生态位分化(图6)。凋落物分解过程中优势微生物的演替遵循如下顺序:首先由快速生长的微生物(如细菌、糖类真菌)定殖,消耗易获得的碳源,其次是具有纤维素分解、半纤维素分解和果胶分解能力的真菌(如子囊菌)和细菌(如放线菌),最后是能够分解木质素和几丁质的担子菌(图6)。具有复杂结构的化合物(例如,木质素、纤维素)分解过程中微生物群落的演替也支持时间生态位分化(图6; 表3)。例如,细菌和真菌群落在添加纤维素后的第一周保持稳定,在纤维素分解的两周内,革兰氏阳性菌的生物量减少了13%,而革兰氏阴性菌和真菌的生物量增加了12-50%。相反,在添加纤维素2个月后观察到相反的模式。这与草原土壤中真菌生物量占总微生物生物量的比例在添加纤维素40天后增加,然后在纤维素耗尽后降低的结果一致。
4 全球变化影响
在全球变化的广泛组成部分中,有5个与土壤过程特别相关,可能影响微生物群落以及细菌和真菌对资源的竞争力:CO2浓度升高、氮沉降、土壤酸化、全球变暖和干旱(图7)。本章还讨论了其他区域气候变化(例如,降水增加、冻土融化、火灾频率增加、土地利用变化、盐碱化和重金属污染)的影响。下文将描述这些影响微生物群落的机制,特别关注真菌和细菌的竞争能力。
理解全球变化过程如何相互作用对于预测细菌-真菌相互作用至关重要。尽管有限的研究探讨了特定区域或生态系统中预期的与气候相关的一系列变化对土壤微生物群落和多样性的影响,但目前还不能得出细菌-真菌竞争相互作用与多因素变化的一般性结论。因此,我们避免呈现这些全球变化成分的潜在交互效应,因为它们所涉及的不确定性非常高。这强调了进行长期多因子实验的必要性,以更好地了解气候变化对细菌-真菌相互作用的影响及其对生物地球化学过程的影响。
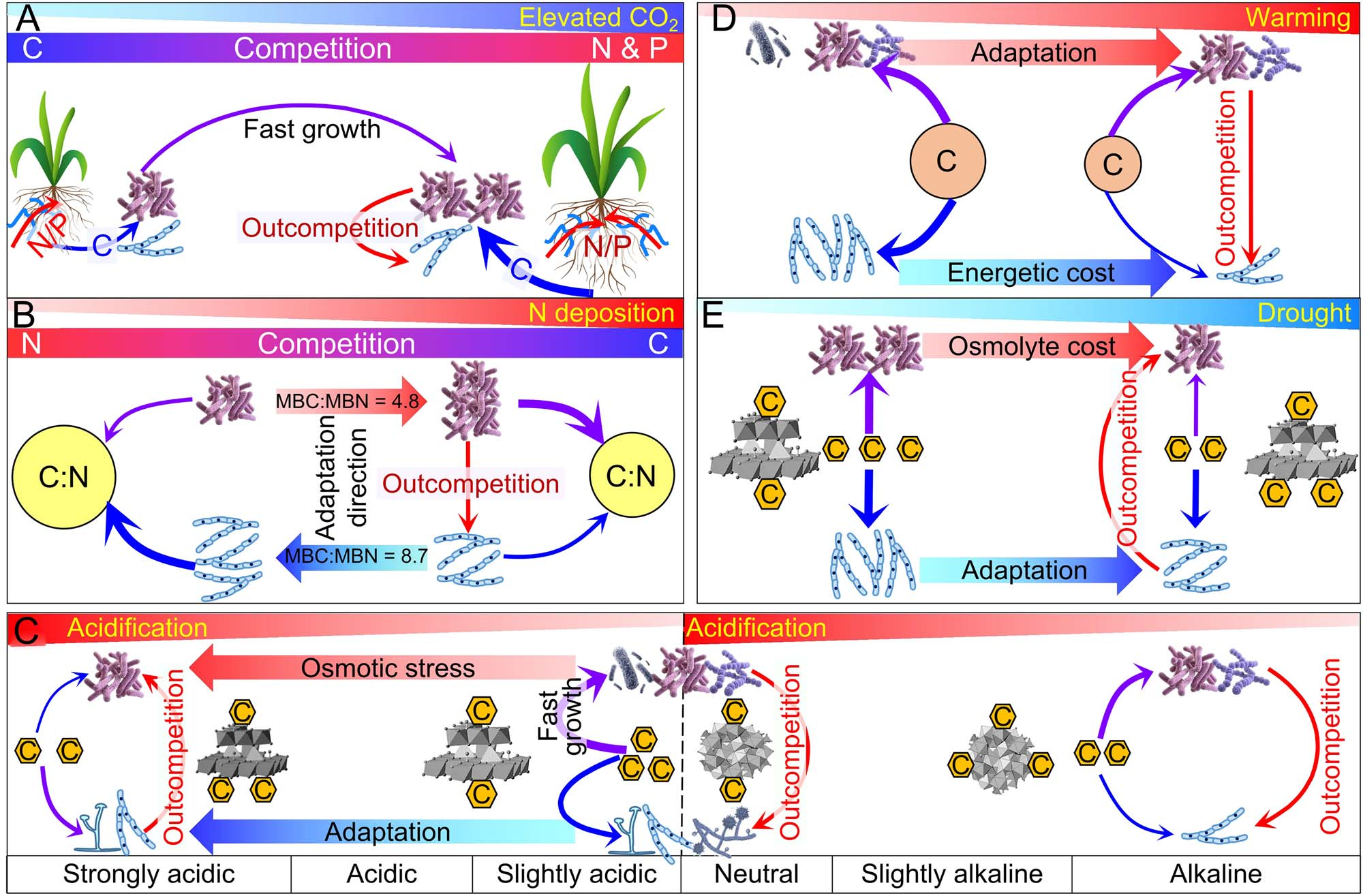
图7 全球变化影响细菌-真菌对碳源和能源的竞争。全球变化的影响包括:CO2浓度升高(a)、氮沉降(b)、土壤酸化(c)、增温(d)和干旱(e)。梯形、箭头和线的厚度表示各个过程的相对强度。
4.1 CO2升高
大气CO2浓度在过去的50年中稳定增加了> 90 ppm,预计到2100年将增加300~600 ppm。在CO2浓度升高的条件下,根系通常会释放更多的C,从而增加微生物活性,尤其是在根际。因此,CO2浓度升高条件下C输入的增加刺激了r-策略菌的生长,主要是细菌。理论上,根系分泌物的增加会降低细菌和真菌对C的竞争强度。然而,C有效性的增加会导致微生物生长的其他营养元素(主要是N和P)受到限制(图7)。这随后引发了细菌和真菌之间对N和P的强烈竞争,这种竞争在养分有限生态系统中尤为激烈(图7)。因此,CO2浓度升高条件下根际沉积增加对竞争的影响在很大程度上取决于N和P的有效性。
在富含N和P的条件下,细菌会迅速利用较高CO2浓度下增加的根际沉积物进行生长,从而提高其生物量。相反,在N和/或P缺乏的条件下,根际沉积增加会刺激微生物分解SOM以获得更多的N和P,即所谓的“priming effect”。这有利于产胞外酶的真菌比细菌更多地利用难分解的有机C。植物对N的低可利用性导致高CO2浓度下根系的菌根化程度更高,从而增加了外生菌根和丛枝菌根群落的丰度。由于细菌对N的需求量至少是真菌的2倍(细菌生物量的C:N比为4.8,而真菌生物量的C:N比为8.7),因此任何N限制对细菌的影响都比真菌强烈,尤其是在CO2浓度升高的条件下。因此,CO2浓度升高降低了对C的竞争,但增加了对N的竞争和真菌的竞争力,因为真菌对N的需求较低(图7)。
4.2 氮沉降
大气氮沉降在20世纪增加了3~5倍。在氮沉降下,细菌对碳源和能源的竞争力预计会增加,因为它们对氮的需求更高(图7)。这一点被所有陆地生态系统荟萃分析评估的氮沉降下真菌/细菌比值的降低所证实。较高的氮有效性减弱了根系与外生菌根和丛枝菌根真菌之间的共生关系,从而降低了这些真菌的丰度。
氮沉降增加会对细菌产生两种截然不同的影响,这取决于碳的可利用性。在富C条件(如根际沉积增加)下,氮有效性的增加刺激了快速生长的革兰氏阴性菌(r-策略菌)的生长。相比之下,随着凋落物的输入,革兰氏阳性菌将主导微生物群落,长期氮沉降后稳定性C积累,以及植物地下C分配减少。在这两种情况下,细菌将比真菌更多地利用C和能源(图7)。然而,氮沉降强烈加速了土壤酸化,这将产生相反的影响。
4.3 酸化
土壤pH对微生物生长和竞争具有重要的调控作用,因为微生物具有各自的最适pH范围。例如,在pH 4~8范围内真菌与细菌生长比的动态变化是细菌竞争性抑制真菌生长的结果。因此,土壤酸化改变了资源的竞争力。引起土壤酸化的最相关过程是施氮肥(例如施用尿素,特别是(NH4)2SO4)。
土壤酸化,特别是那些已经酸化的土壤,加剧了对C和能源的竞争,原因有二:(i)由于酶和/或底物在倍半氧化物表面的吸附,导致胞外酶活性和/或底物可用性大幅度降低;(ii)增加碳和能源分配以缓解酸性胁迫(图7)。例如,当pH从4.5下降到3.8时,酿酒酵母(Saccharomyces cerevisiae)的ATP消耗增加了近100倍,而ATP产量没有变化。酸化提高了真菌在酸性土壤中的竞争力,因为真菌具有更高的渗透胁迫耐受能力(图7),并且由于细菌产生的胞外酶的最适pH在高(中性至碱性)pH下表现最好,而真菌胞外酶在低(酸性)pH下表现最好。
4.4 全球气候变暖
温度是微生物代谢、生物量、群落组成、群落演替和物种间相互作用的决定因素之一。这是因为微生物物种在土壤中对温度变化的适应性和适合度差异很大。值得注意的是,增温有许多间接影响,因为它改变了在选择微生物物种方面发挥作用的其他因素(例如,土壤湿度、植被类型和生产力、时间)。一项长期(长达26年)研究提出了SOM分解的四个一般阶段以及与土壤增温相关的微生物机制:通过呼吸作用快速损失C;活性C库耗竭后,微生物群落发生重组,真菌生物量降低,革兰氏阳性菌丰度增加;向具有较低真菌优势和真菌/细菌比值的更多样化、寡营养微生物群落转变;以及稳定性C库和微生物生物量的减少。
尽管增温降低了微生物生物量和多样性,但真菌生物量和多样性的降低幅度大于细菌(图7)。例如,7年升温+3℃使草地土壤细菌和真菌丰富度分别降低了9.6%和14.5%。增温增加了细菌的优势度,因为真菌比细菌对温度更敏感,而最适温度(即具有最大活性的温度)和最高温度敏感点(即活性变化最大的点)则相反。这意味着细菌对增温的适应性更好(图7)。随着气候变暖,细菌群落向更嗜热的类群转变,而真菌功能群不太可能发生改变。这得到了不稳定碳分解种群和基因增加的支持,而稳定性碳分解的种群和基因在9年2℃增温下保持稳定。因此,细菌有望成为增温条件下C源和能源的更好竞争者(图7)。
增温可能会加剧竞争,因为所有的微生物都需要更多的C和能量资源来满足其随着温度升高而增加的代谢需求。真菌(-14 kJ mol-1 C ℃-1)的热容是细菌(-5 kJ mol-1 C ℃-1)的3倍。因此,真菌必须分配更多的能量用于生物量的维持和生长。例如,假设土壤微生物以葡萄糖为底物的能量利用效率为0.32,微生物将1 mol葡萄糖完全氧化为6 mol CO2并产生38 mol ATP (-30.5 kJ mol-1),那么在温度升高1℃的情况下,通过卡尔文循环,细菌比真菌更多地将68 mmol C转化为其生物量。因此,土壤增温削弱了真菌的竞争力。
4.5 干旱
干旱增加了对C源和能源的竞争强度(图7)。首先,当土壤变得干燥时,微生物必须通过积累溶质(渗透调节物质)来平衡土壤中的渗透条件,以保持细胞内的水分。缓解干旱胁迫的另一种策略是产生胞外聚合物(例如,多糖、蛋白质等),这些物质可以充当海绵延缓干燥。渗透调节物质的积累是一个能量昂贵的过程,需要C,从而减少了用于生物量维持和繁殖的C和能量分配。其次,由于有机化合物在金属氧化物上的吸附增加,干旱可能会降低碳的有效性。这可以达到离子溶液中产物的溶解度,并导致胶体溶液的排斥电荷崩塌。相比之下,干旱条件下的离子强度比最佳水分条件下高出9个数量级,这可能导致C从矿物中解吸。然而,干旱条件下土壤水分的减少极大地限制了碳的扩散,从而减少了微生物,尤其是细菌对C的可利用性。第三,干旱降低了各种水解酶的活性,从而降低了SOM的分解速率。
对水分限制抵抗力较强的真菌在干旱条件下能够比细菌更多地竞争碳源和能源(图7),这主要是因为真菌菌丝可以桥接空间上离散的资源,也因为真菌产生的有效胞外酶(特别是氧化酶)可以分解复杂的有机化合物。
4.6 其他全球变化因素
降水增加主要发生在潮湿的热带和北部地区。尽管降水量的增加对真菌/细菌的比值没有直接影响,但随着土壤含水量的增加,尤其是当土壤初始含水量较低时,细菌可能会在所有资源方面胜过真菌。同样,随着永久冻土融化并使孔隙充满水,细菌C和能量通道可能主导北极的SOC动态。虽然真菌和细菌对重金属污染的响应没有一致的结论,但前者普遍具有较高的抗性。因此,真菌可能胜过细菌,特别是随着污染程度和持续时间的增加。
与上述全球变化因素不同,总结它们对细菌-真菌竞争生态系统中争夺碳和能量的影响是具有挑战性的,因为这些影响取决于多种因素及其相互作用。例如,细菌和真菌丰度的变化取决于土地利用方式。农田和耕作面积的增加会显著降低微生物生物量,尤其是真菌生物量,从而导致农业土壤中细菌的优势地位。稻田水分管理的逐渐改变导致更好的土壤通气性,并向真菌群落和革兰氏阳性菌转变。
野火频率增加对这种竞争的影响取决于火灾的严重程度和持续时间、土壤恢复力和环境条件,两者都对热敏感。然而,野火后表面残留的持久性致热产物以及土壤和有机质疏水性的增加将导致真菌群落的强烈转变。
尽管真菌通常具有更强的应对渗透胁迫的能力,但这两个类群对盐渍化的响应模式具有生境或环境特异性。盐渍化,尤其是在低盐土壤中,可能会加速对盐胁迫抵抗能力较差的细菌的生物量损失。
5 细菌和真菌共存
土壤微生物多样性对陆地生态系统的功能、稳定性和健康至关重要。纯竞争或合作群落的稳定性弱于具有复杂相互作用的群落。尽管细菌和真菌在土壤中强烈竞争资源,但它们总是共存于相同的生态位,并形成了各种协同相互作用(如交叉喂养、互利共栖、共生等)。这种共存有助于这两个群体提高其能够取食的底物的质量和数量,并扩大其空间生态位空间。这也增加了它们对环境变化的抵抗力。
代谢依赖性是这种共存和微生物群落稳定性的主要驱动因素。虽然两个群体从根本上竞争资源,但来自一个生物体的排泄产物可能是另一个生物体偏好的C、能量和营养来源,导致交叉喂养(图3)。由于取食偏好的巨大差异,这种交叉喂养相互作用在土壤中是复杂而普遍的(图4)。在交叉喂养中,一个种群释放的代谢物增加了另一个种群的底物数量。
交叉喂养的细菌和真菌更能抵御环境胁迫,尤其是养分限制下。真菌可能受益于细菌的存在,特别是在获取有机氮和去除生长抑制剂方面。担子菌和丛枝菌根缺乏直接获取有机氮的高效胞外酶。在固氮菌与担子菌之间的交叉喂养相互作用中,细菌不断向担子菌提供N,作为交换,细菌利用真菌胞外酶释放的低聚物作为C、能量和其他营养物质(如P)。交叉喂养相互作用在这种共存中至关重要,特别是当一个种群消耗另一个种群释放的有毒代谢物,从而促进代谢物产生种群的生长时。
真菌的存在有助于细菌扩大生态位空间。Pseudomonas aeruginosa PAO-rfp在有青霉菌菌丝时的总扩展半径是没有菌丝时的1.4倍证实了这一点。这主要是由于真菌创造了一个微水物理环境,增加了细菌的运动性,从而使细菌能够定殖在未被占据的生态位中。这种正效应强烈调节着土壤中细菌的多样性和功能。例如,丛枝菌根真菌的根外菌丝将溶磷细菌输送到有机磷斑块,从而加速有机磷矿化。这种共存增加了对化感物质的抗性,从而缓解了此类物质对种群扩张的负面影响。在Burkholderia terrae BS001与非木材腐朽真菌之间的互利共生关系中,Burkholderia terrae BS001通过拮抗剂(如环己酰亚胺,拮抗细菌的代谢产物)的吸附或解毒作用来保护真菌。反过来,细菌通过获得菌丝释放的有机化合物和营养物质而获益。
复杂物种的网络共存增加了微生物群落对环境变化的抵抗能力。例如,尽管增温条件下微生物OTU数量比对照减少了12%,但由此产生的正相关比例较对照增加了43%。这表明微生物群落适应增温的主要策略是促进类群间的合作行为。
结论与未来展望
土壤中细菌和真菌对C、能量和养分的竞争是普遍存在的。这使得这种相互作用的后果及其对生物地球化学过程的影响对土壤功能至关重要。本文证明,细菌在利用简单有机化合物作为底物时效率高出1.4-5倍,而真菌在利用复杂和持久性化合物时效率高出1.1-4.1倍。这主要是因为细菌通过更简单的细胞膜结构更快速地结合小分子有机物。相比之下,真菌产生非常高效的胞外酶来分解复杂的化合物,使其能够在分解复杂化合物过程中缓慢地获得C和能量。
上述强烈的剥削性竞争导致了化学、空间和时间生态位分化。具体而言,细菌在利用亲水性化合物(化学生态位)方面胜过真菌,在非根际土壤、深层土壤、大团聚体、小孔隙(空间生态位)和缺氧位置(化学生态位)中主导C的利用,并且在植物凋落物分解的早期阶段主导C的利用。相反,有效的胞外酶有助于真菌在疏水性和持久性(如木质素和几丁质)化合物以及富含复杂化合物的生态位中胜过细菌。根系定殖增加了真菌对根际土壤中C和能量来源的竞争力。
未来需要解决的重要问题:(i)细菌和真菌对碳和能量的需求和投资有何差异,使它们成为成功的竞争者?(ii)在细菌和真菌之间竞争加剧的情况下,生态位分化的机制及其后果是什么?(iii)在空间和时间尺度上,这种对C、能量和养分的竞争的机制及其后果是什么?(iv)各种资源的变化(如全球变化因素)如何改变细菌和真菌的竞争力及其在土壤中发挥作用的后果?
原文链接:
https://academic.oup.com/ismej/article/18/1/wrae073/7660942#453602202
-
点赞 (0人)
- 收藏 (0人)

