
Molecular Ecology最新研究:土壤微生物的子群落构建机制及其与各微生物类群生态属性和功能特征的关系
- 看不见的线
- 5771
- 2024-06-06 10:00:11
- 文章来源:土壤宏组学
原文信息
原文标题:Soil microbial subcommunity assembly mechanisms are highly variable and intimately linked to their ecological and functional traits
发表期刊:Molecular Ecology
影响因子:4.9
期刊分区:中科院一区
发表时间:2024.02.29
共同一作:范秋萍;刘开放
通讯作者:车荣晓
第一单位:云南大学 国际河流与生态安全研究院
论文链接:
https://doi.org/10.1111/mec.17302
编 译:刘开放 云南大学国际河流与生态安全研究院
摘要
揭示土壤微生物群落构建机制是分子生态学研究的重要目标之一。尽管对整体微生物群落构建机制的研究日益增多,但对不同微生物类群子群落构建机制的认识仍十分有限。本研究采集了来自云南地区100多个地点的土壤样品。基于扩增子高通量测序和iCAMP分析,本研究揭示了各微生物类群的子群落构建机制。结果显示,扩散限制和同质选择是该地区土壤微生物群落构建的主要驱动因素。不同土壤微生物类群的子群落构建机制存在较高的异质性。例如,同质选择对古菌子群落构建的贡献率为70%,但对放线菌门、芽单胞菌门和浮霉菌门子群落构建的贡献仅10%左右。具有较高出现频率、平均相对多度和网络关联度,以及更宽生态位的微生物子群落构建往往更受均质扩散和漂变的影响,但受到异质选择和扩散限制的影响较小。不同功能群的子群落构建机制也存在较大的变异。值得注意的是,固氮菌、硝化菌、腐生微生物和一些土传病原菌的子群落构建主要受同质选择控制,而反硝化菌和真菌病原菌的子群落构建主要受漂变等随机过程的影响。这些发现为我们理解土壤微生物多样性维持机制提供了新的见解,而本研究的分析流程对未来的研究也具有重要价值。
1. 引言
解析微生物群落构建机制一直是生态学研究的基本目标。一系列假说已被提出,以解释微生物群落构建机制。生态位假说认为,微生物群落构建主要受选择等确定性过程驱动。相反,中性理论认为,微生物群落的构建主要受到随机出生、死亡、定殖、灭绝和物种形成的驱动。最近提出的过程理论则认为,微生物群落构建主要由四个生态过程决定:选择、扩散、漂变和多样化。所有上述生态过程通常作用于微生物群落的精细分类水平而非整体分类水平。因此,具有不同生态属性的土壤微生物子群落构建机制也不同。例如,选择对由细胞大小较小且相对多度较高的类群组成的土壤微生物子群落构建更为重要。然而,这些差异无法通过对微生物群落整体构建机制的分析来识别。因此,探索微生物谱系在细微分类水平上的子群落构建机制可以为我们提供对微生物多样性维持机制的关键见解。此外,了解功能群的子群落构建机制可以指导土壤微生物组操纵,改善土壤健康。例如,如果植物促生菌的子群落构建主要受到扩散限制的控制,通过有针对性的接种可以潜在地增加它们的多度。总体而言,解析微生物子群落构建机制具有理论和实际意义。
随着βNTI-RCBray等间接推算方法的发展和广泛应用,我们对微生物群落构建机制的关注日益增加。多项研究已经确定了限制扩散、同质选择、异质选择、均质扩散和漂变等过程在微生物群落构建中的作用。iCAMP是一种最近开发出的基于系统发育分箱的零模型算法,主要用于揭示微生物子群落的构建机制。基于该方法,一些研究解析了土壤真核生物的子群落构建机制,以及它们与细胞大小和生态位宽度之间的关系。然而,对土壤原核生物和真菌子群落在更精细的分类水平上的构建机制及其决定因素仍知之甚少。
不同土壤微生物类群和功能群可能受到不同子群落构建机制的影响,这是由它们特定的生态属性和功能特征所决定的。首先,不同微生物类群的生态位宽度差异很大。可以适应广泛环境条件的微生物类群被归类为泛化种,它们通常对环境变化具有抗性。相反,特化种是生态位较窄的微生物类群,它们通常对环境条件的变化比较敏感。因此,环境选择对泛化种和特化种的子群落构建的贡献可能会有很大不同。其次,微生物群落是由少量丰富种和大量稀有种共存而形成的。稀有和丰富微生物子群落的构建机制存在很大的差异。第三,不同微生物类群之间存在竞争、合作和拮抗等复杂相互作用,这也会显著影响微生物类群的子群落构建。最后,不同微生物类群的传播能力也可能存在很大差异。总而言之,不同微生物类群的子群落构建机制可能会因其生态属性和功能特征的不同而表现出高度异质性。
本研究旨在揭示云南省不同土壤微生物谱系和功能群的区域尺度子群落构建机制,并解析其主要影响因素。我们收集了云南省的土壤样品,通过对原核生物16S rRNA基因和真菌内转录间隔子(ITS)进行高通量测序分析土壤微生物多样性。利用iCAMP算法确定了微生物类群的子群落构建机制。在此基础上,结合iCAMP、FUNGuild和FAPROTAX,建立了新分析流程,确定了微生物功能群的子群落构建机制。基于以上发现,我们提出了两个假设:不同微生物群的子群落构建机制差异显著;不同微生物群之间子群落构建机制的差异主要取决于它们的生态属性和功能特征。
2. 材料与方法
2.1 研究地点和土壤采样
研究地点位于中国西南部典型高原山地——云南省。该地区的环境多样性丰富,具有亚热带季风、热带季风和高原山地气候等多种气候类型。年均温和年降水量的范围分别为4.7~23.3℃和613~1733毫米(表S1)。云南省的多样化环境也使其成为全球生物多样性热点。
土壤样品于2019年9月至10月在云南省的117个样点采集,采样点大致均匀分布在研究区域内(图S1和表S1)。大多数研究地点表现出多样化的土地利用模式,包括森林、农田和草地。为减少由这些土地利用模式所导致的潜在采样偏差,所有土壤均采集自玉米地。此外,还收集了30个相邻森林的土壤样品,以进一步明确不同土地利用模式对土壤微生物群落构建的潜在影响(图S1和S2)。在每个采样点,设置了间隔超过10米的五个亚采样点。使用土钻采集了0~10厘米深度的土壤样品。然后,每个采样点的亚采样点采集的土壤样品,过2毫米筛后彻底混匀。随后,每份混合土样被分成三份。具体而言,约10克土壤存放在液氮中,并在实验室中以-80°C保存以供DNA提取。约200克土壤在室温下风干,以确定pH值、质地,以及总碳、总氮、速效磷和速效钾的含量。此外,保留了200克新鲜土壤在4°C下测定铵态氮、硝态氮和含水量。通过GPS系统获取了每个采样点的经度、纬度和海拔信息,并从WorldClim数据库(www.worldclim.org)收集了MAT和MAP数据。
2.2 土壤理化性质分析
土壤在105℃下干燥24小时来测量含水量。土壤pH是使用1:2.5的土壤-水比率的pH计检测的。土壤总碳和总氮的含量使用自动元素分析仪测量。基于分光光度法测定土壤中的铵态氮和硝态氮浓度。分别使用Olsen和Bray法测定了碱性和酸性土壤中的速效磷含量。采用冷硝酸法提取速效钾,并利用流动注射仪器测定速效钾含量。使用激光颗粒分析仪测量土壤质地。根据前期研究提出的标准,将土壤颗粒分为粘粒(0~2 μm)、粉粒(2~53 μm)和砂粒(53~2000 μm)。
2.3扩增子高通量测序
使用DNeasy PowerSoil Kit从0.30克冻土中提取DNA,按照制造商的说明进行。使用Nanodrop 2000超微量分光光度计确定DNA提取物的质量和浓度。原核16S rRNA基因的V4–V5区域用引物515F(5’-GTG CCA GCM GCC GCG GTA A-3’)和909R(5’-CCC CGY CAA TTC MTT TRA GT-3’)扩增。真菌ITS的通用引物使用gITS7(5’-GTG ART CAT CGA RTC TTT G-3’)和ITS4(5’-TCC TCC GCT TAT TGA TAT GC-3’)。50 μL的PCR体系包含5 μL的ExTaq缓冲液、4 μL的dNTP混合液、1 μL的前段引物、1 μL的后端引物、0.25 μL的ExTaq、37.75 μL的无核酸PCR水和1 μL的模板DNA。所有的PCR都是在Bio-Rad S1000 PCR系统上进行的。PCR循环从94℃的初始变性开始,持续5分钟,然后进行30个循环,包括94℃的变性,持续30秒,56℃的退火,持续30秒,72℃的延伸,持续30秒,以及72℃的最终延伸,持续10分钟。根据我们的初步测试,本研究中16S rRNA基因和ITS的最佳PCR条件是相同的。PCR产物使用Axyprep DNA凝胶提取试剂盒纯化。库构建是根据NEBNext® Ultra™ II DNA文库制备套件的标准程序进行的,并采用Illumina Nova 6000平台进行配对末端测序。
2.4 生物信息学分析
16S rRNA基因和ITS序列数据根据UPARSE与USEARCH(v11)进行分析。使用UNOISE3非聚类去噪算法获得了zOTUs的代表序列。删掉了少于9个序列的zOTUs。然后,使用USEARCH的“otutab”命令基于97%相似性阈值生成了zOTU表。随后,使用QIIME2对代表序列进行了与原核生物和真菌分别相对应的Silva138和UNITE 8.2数据库的系统发育信息的比对,以获得分类注释信息。使用vegan软件包中将每个样本中的原核生物和真菌序列数分别稀疏至36901和77314。使用vegan软件包中的diversity函数计算了微生物丰富度。根据核糖体RNA操纵子数据库(rrnDB v5.8)确定了每个OTU的rRNA操纵子拷贝数。
16S rRNA基因和ITS的系统树分别采用了最大似然法在QIIME2中构建。最近,人们对ITS系统树的潜在局限性引起了高度关注。然而,最近的研究表明,基于原始ITS树和从UNITE数据库导出的调整树的iCAMP分析结果几乎相同。一个可能的解释是ITS树的问题主要是由长距离的系统距离引起的,而iCAMP主要集中在高度相关的分类单元上。为了减少调整系统树引入的潜在偏倚,我们采用了相同的方法来构建16S rRNA基因和ITS的系统树进行接下来的分析。
2.5 土壤微生物群落子群落构建机制分析
使用iCAMP算法检验了群落和子群落构建机制。首先,根据它们的系统发育关系将zOTUs分为不同的箱。简而言之,系统发育树在距离根部的某个特定距离处被截断(尽可能短),距离设置为默认值0.2,所有剩余的连接低于此阈值。在截断点之后,源自同一祖先的分类单元被分组到相同的箱中。然后,将每个小箱合并到包含最近亲属的箱中。重复此过程,直到所有合并的箱中都有超过24个zOTUs。随后,基于零模型分析,计算了每对样品之间所有箱中的βNRI和修改后的Raup-Crick指标。通常,每个箱中的所有分类单元都具有密切的系统发育关系,因此在这种情况下,βNRI比βNTI表现优异,因此iCAMP采用了βNRI而不是βNTI。对于每个分箱,具有βNRI < -1.96的样品对之间的转变被认为是受同质选择控制的,而在具有βNRI > 1.96的样品对之间的转变则被认为是受异质选择控制的。使用RC指标对具有|βNRI| ≤ 1.96的成对比较进行了分区。在结果中,具有|βNRI| ≤ 1.96和RC <-0.95的转变被控制为均质扩散,而具有|βNRI| ≤ 1.96和RC > 0.95的转变被控制为扩散限制。具有|βNRI| ≤ 1.96和|RC| ≤ 0.95的转变主要受到漂变的驱动。在iCAMP分析中,漂变,被定义为既不受选择也不受扩散控制的转变,通常包括漂变、多样化、弱选择或弱扩散。然后,将所有分箱中的各个过程的贡献按照每个箱的相对多度进行加权,汇总以估计在一组群落的构建中各个过程的相对重要性。使用icamp.cate函数评估了不同分类水平上子群落的各个过程的相对重要性。最后,基于所有微生物子群落的构建机制的加权总和确定了整体微生物群落的构建机制。
使用FAPROTAX(v1.2.4)和FUNGuild(v1.2)进行了原核生物和真菌的功能预测。真菌功能群的子群落构建机制直接基于FUNGuild的输出文件进行了分析。但是,对于原核生物功能群,根据本研究所开发的R脚本,从faprotax_report文件中的功能注释添加到了基于此的原核生物zOTU表。然后,使用icamp.cate函数确定了预测的原核生物功能组的群落构建过程。这些R脚本和示例注释可在GitHub(https://github.com/LKF98)上获取。
2.6 生态属性的确定
生态位宽度表示特定微生物分类单元可以适应的环境范围。在本研究中,使用“spaa”软件包中的Levin方法计算了生态位宽度。平均相对多度和出现频率反映了每个微生物分类单元的稀有度和出现率,分别用“base”包在R中计算。在zOTU级别确定了生态位宽度、平均相对多度和出现频率。使用“car”包计算了这些生态属性之间的多重共线性,分析显示,所有生态属性的VIF值均低于4,表明它们之间没有显著的多重共线性问题。网络度表示与节点连接的边的数量,并反映了微生物分类单元之间的共存模式。基于SparCC软件包进行了每个箱之间的Spearman相关性计算,仅包括ρ ≥ 0.5和P < 0.01的强相关性。使用“igraph”软件包确定了网络属性,如网络度。所有生态属性都是在R(4.1.0)中进行的。
2.7 统计分析
使用ArcGIS(v 10.2)绘制土壤性质、微生物群落组成和丰富度地图。基于Bray-Curtis距离进行非度量多维尺度分析(NMDS),可视化了原核生物和真菌群落结构的差异。基于欧几里德距离进行了土壤性质的非度量多维尺度分析(NMDS)。通过envfit函数、Mantel检验和CCA分析了环境因子与微生物群落结构之间的关系。Mantel检验还用于确定群落构建过程与环境变量之间的相关性,并使用“corrplot”包进行可视化。在“vegan”和“ecodist”包中,使用999次置换确定了统计显著性。利用Spearman相关性检验确定了亚群构建机制与微生物分类属性(即生态位宽度、相对多度、出现频率、网络度和rRNA操纵子拷贝数)之间的关系。所有上述统计分析均在R(4.1.0)中进行。
3. 结果
3.1 土壤性质、微生物多样性和群落特征
研究地点的土壤性质差异很大,表明种植玉米未能消除该地区土壤固有的异质性(表S1;图S3和图S4)。云南的土壤微生物也具有很高的多样性,原核生物和真菌的总zOTU数量分别达到了27000和34000。然而,不同研究地点的土壤微生物多样性差异很大(图1)。例如,所有样本点中最低的原核生物和真菌丰富度仅为3120和953,而最高的则分别超过9000和3000(图1a和1b)。就微生物群落组成而言,酸杆菌门、变形菌门、拟杆菌门、泉古菌门、绿弯菌门和浮霉菌门是主要的原核生物门,它们占原核生物群落成员的90%以上(图1c)。真菌门的主要门类是子囊菌门(54.6%)和担子菌门(8.39%),其他门类的相对多度通常相当低(图1d)。
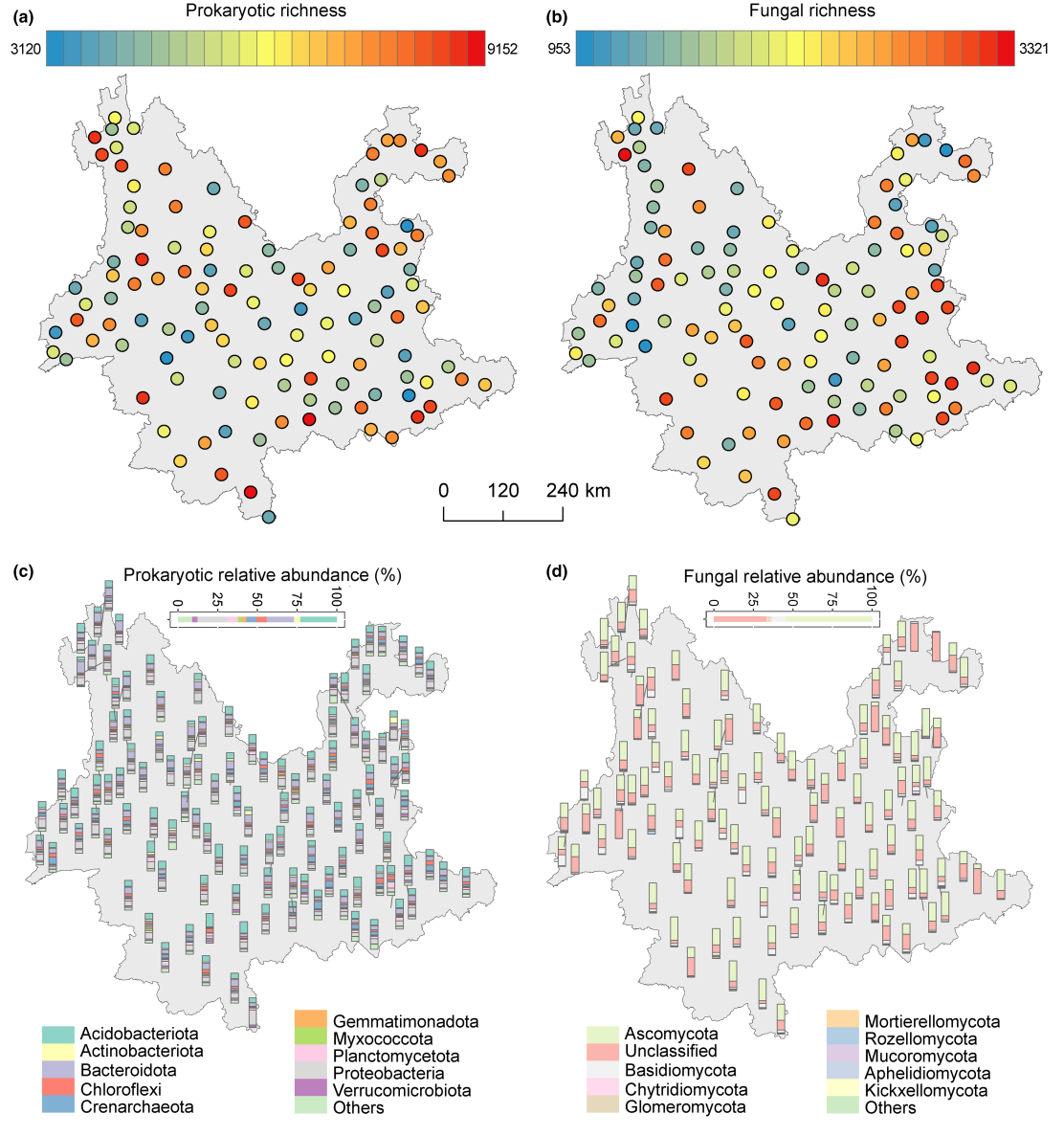
多个环境因素与土壤微生物群落组成和丰富度显著相关。具体来说,土壤含水量和年均温与原核生物丰富度显著相关,而pH、总氮含量和速效钾含量与真菌丰富度显著相关(表S2)。此外,土壤微生物群落组成与年均温、pH、年降水量、海拔和含水量呈显著相关(图S5)。在这些因素中,pH值和年均温分别是土壤原核生物和真菌群落结构的主要预测因子(图S5)。然而,在本研究中所检测的环境因素仅能解释微生物群落特征总变异的不到5%(图S6)。我们还检验了每个分类单元的子群落特征与环境因素之间的相关性。超过一半的分类单元的子群落特征与环境因素呈现出显著但较弱的相关性(表S3和S4)。
3.2 土壤微生物群落和子群落构建机制
iCAMP分析显示土壤微生物群落构建主要受到扩散限制的控制,其对土壤原核生物和真菌群落构建的贡献分别达到40.5%和53.4%(图2a和2b)。此外,同质选择和漂变也对土壤微生物群落的构建至关重要(图2a和2b)。然而,土壤真菌和原核生物的群落构建机制存在一些差异(图2a和2b)。具体来说,与真菌相比,土壤原核生物群落的构建对扩散限制和异质选择的影响较小,但更依赖于同质选择、均质扩散和漂变(图2a和2b)。有趣的是,研究区内农田和森林中观察到了相似的土壤微生物群落构建模式(图S2)。
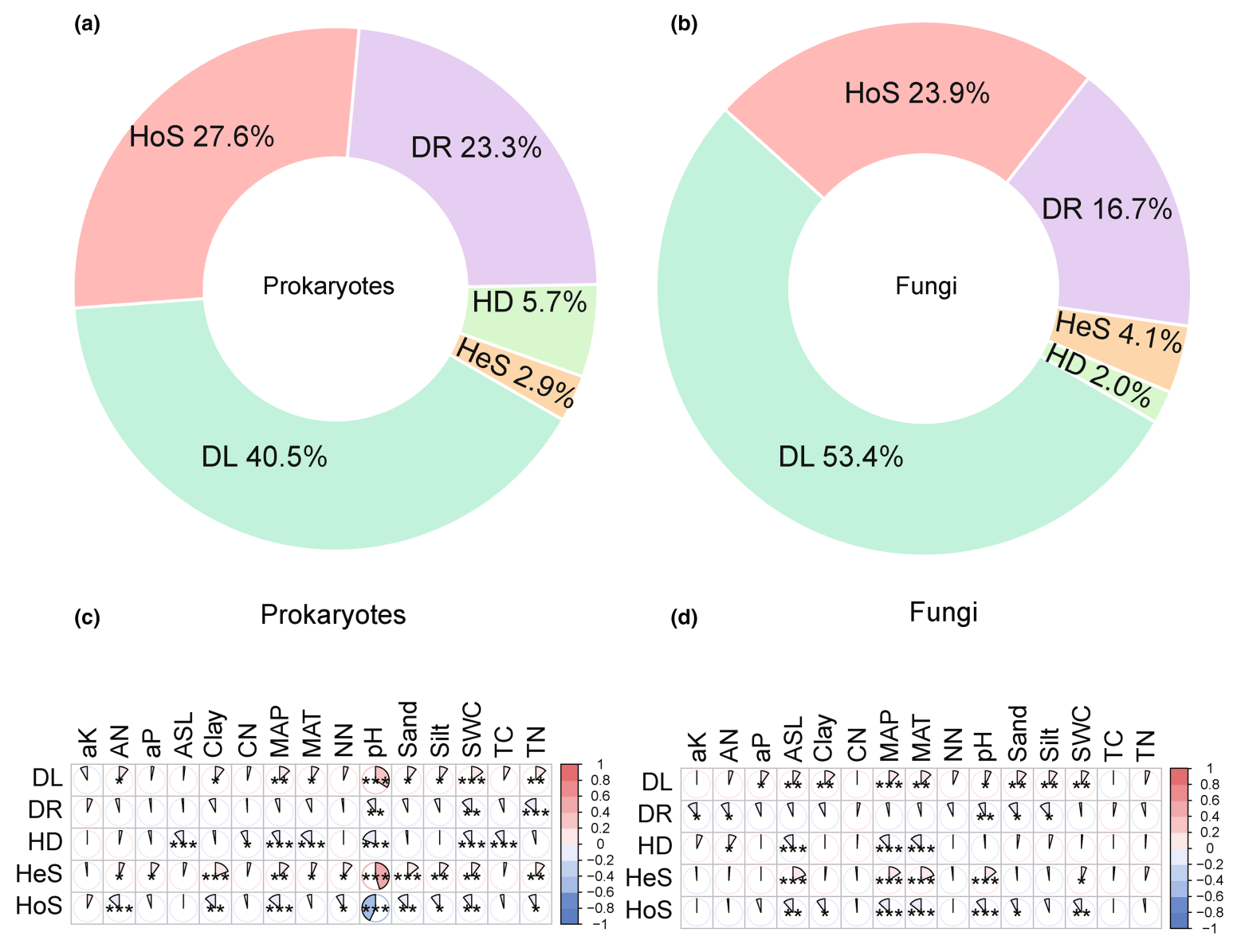
生态过程对微生物群落构建的相对贡献与总体环境距离呈显著相关(图S7)。具体来说,环境差异越大,异质选择和扩散限制的贡献通常越高,而同质选择、均质扩散和漂变的贡献通常越低(图S7)。此外,还观察到这些关系在原核生物和真菌之间是类似的,但对于原核生物而言,相关性明显强于真菌(图S7)。至于具体的环境因素,土壤原核生物群落与pH值呈强相关(图2c)。具体来说,不同生境之间pH的差异越大,土壤原核生物群落构建中同质选择效应越弱,而异质选择和扩散限制效应越强(图2c)。相反,真菌群落构建主要与年均温、年降水量和海拔相关(图2d)。
不同微生物门的子群落构建机制存在显著差异(图3)。对于原核生物而言,同质选择对泉古菌门子群落构建的贡献约为70%,但对放线菌门、芽单胞菌门和浮霉菌门子群落构建的贡献仅约为10%(图3a)。对于真菌而言,漂变和扩散限制主要控制了Aphelidiomycota、罗兹菌门和毛霉门子群落的构建,而被孢霉门子群落的构建主要由同质选择控制(图3b)。同一门类或属的不同级别在同一门类中的子群落构建机制可能存在很大差异(图3c和3d)。例如,对于酸杆菌门而言,维氏菌纲子群落的构建主要受同质选择控制,而酸杆菌纲子群落的构建主要受扩散限制控制(图3c)。
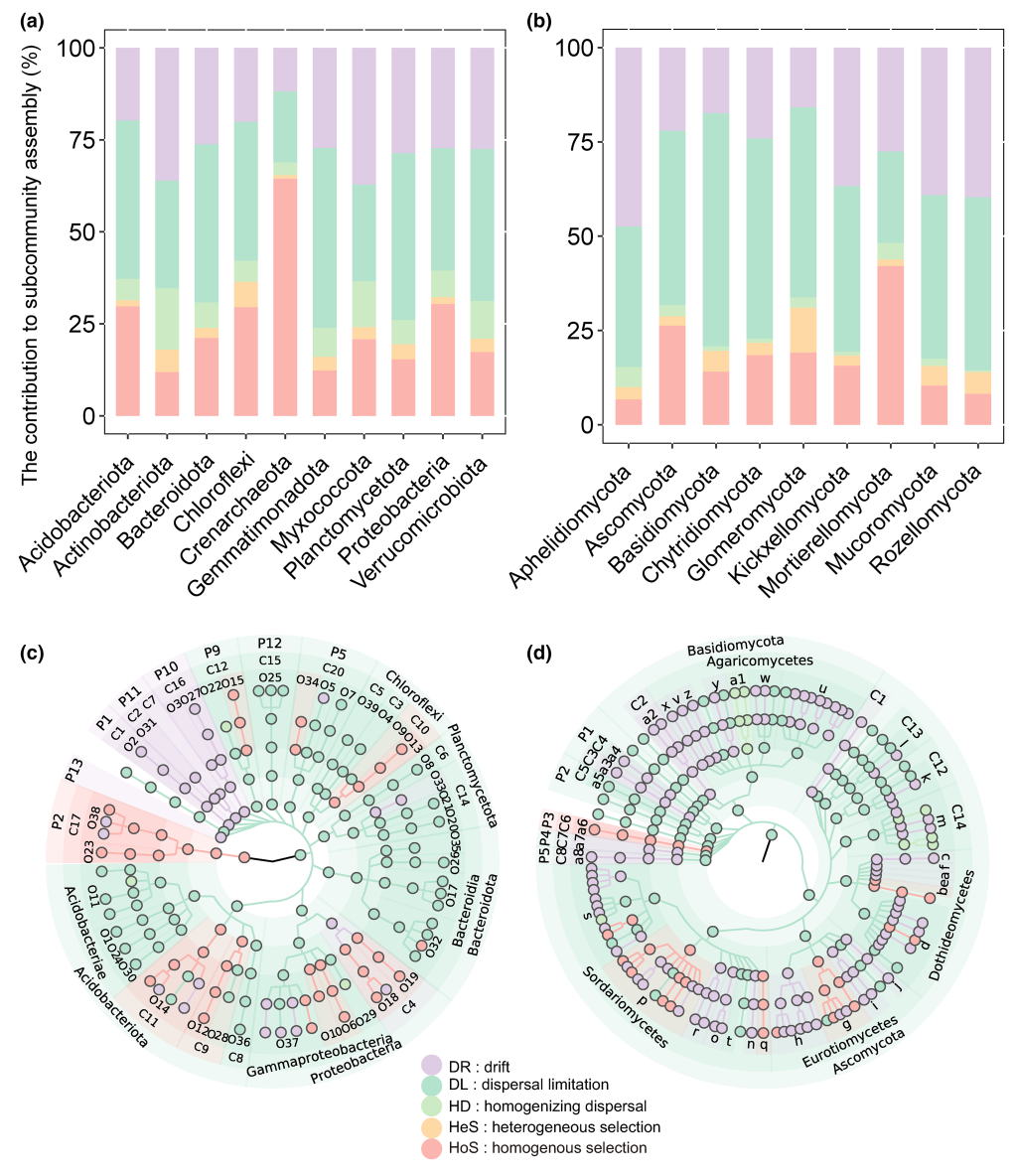
原核生物和真菌分别被划分为495和544个系统发育分箱(图4,表S3和S4)。不同分类单元的子群落构建存在显著差异(图4,表S3和S4)。同质选择对30个原核生物分类单元的构建贡献超过50%,对某些属于变形菌门、酸杆菌门、泉古菌门、拟杆菌门和粘球菌门的分类单元甚至超过90%。异质选择对微生物子群落构建的影响较小,大多数系统发育分类单元的构建贡献不到10%(图4a和表S3)。异质选择对泉古菌的子群落构建的最大贡献为38.7%,其次是绿弯菌的35.9%(图4a和表S3)。相对于其他生态过程,均质扩散对每个原核生物分类单元的构建贡献最小,几乎所有分类单元的贡献都低于20%(图4a和表S3)。超过200个原核生物分类单元的构建中,扩散限制的贡献超过50%,甚至在某些属于拟杆菌门、粘球菌门、酸杆菌门和绿弯菌门的分类单元中达到80%(图4a和表S3)。漂变对43个原核生物分类单元的构建贡献达到50%或更高,对属于硝化细菌门、拟杆菌门、粘球菌门和绿弯菌门的分类单元的贡献超过70%(图4a和表S3)。
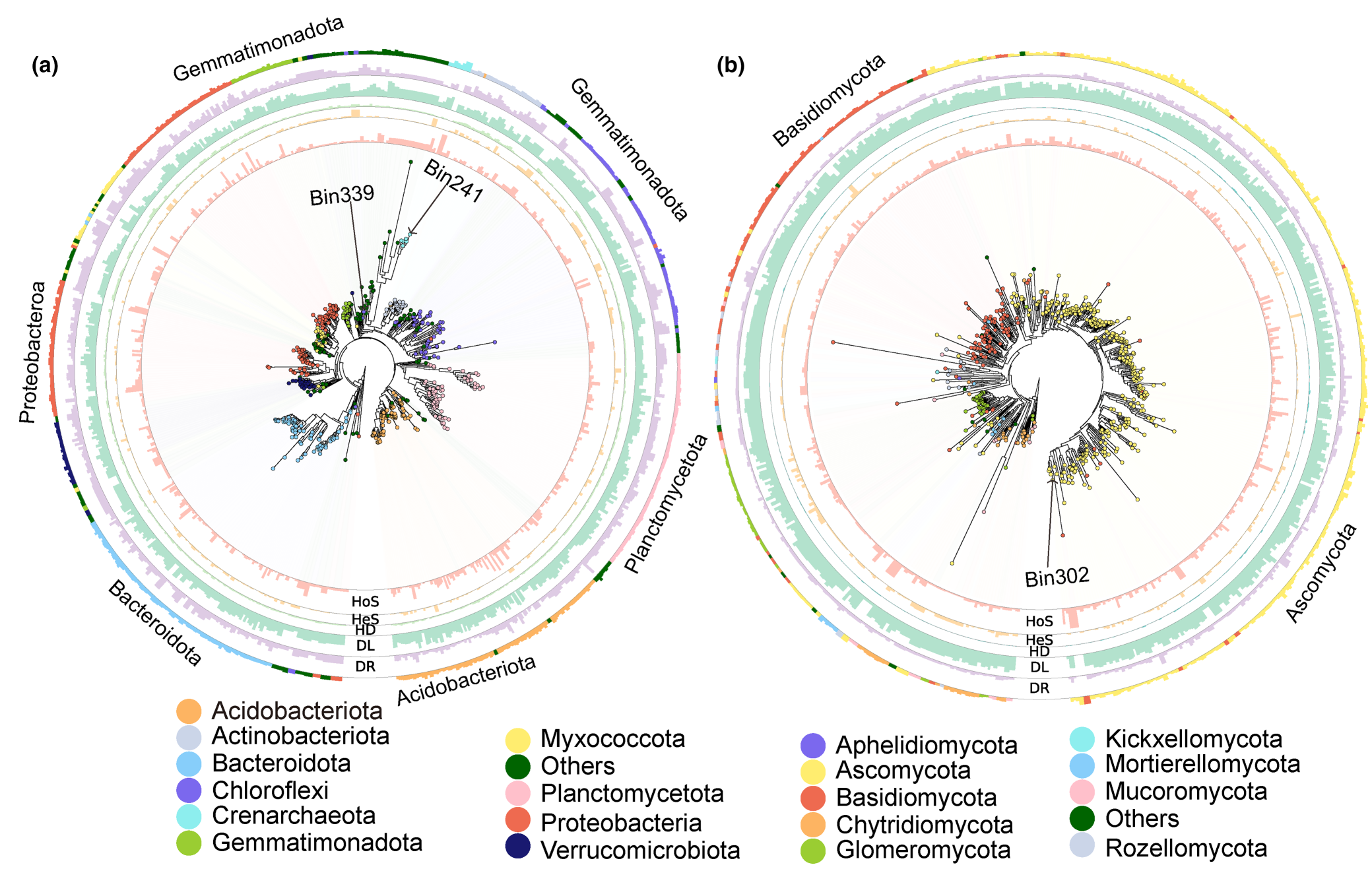
26个真菌分类单元的子群落构建中,同质选择的贡献超过50%,其中子囊菌门的bin302的贡献最大,为97.6%(图4b和表S4)。异质选择对每个真菌分类单元的子群落构建的贡献有很大差异,只有8个真菌分类单元的贡献超过30%,特别是担子菌门和子囊菌门的一些分类单元的异质选择贡献达到50%(图4b和表S4)。与此相反,均质扩散对真菌子群落构建的影响几乎可以忽略,没有任何贡献超过10%(图4b和表S4)。相比之下,425个真菌分类单元的子群落构建中,扩散限制的贡献超过50%,甚至对属于球囊菌门和子囊菌门的一些分类单元的贡献达到90%(图4b和表S4)。此外,漂变对4个真菌分类单元的子群落构建的贡献超过50%(图4b和表S4)。
3.3 不同土壤微生物类群子群落构建机制的潜在驱动因素
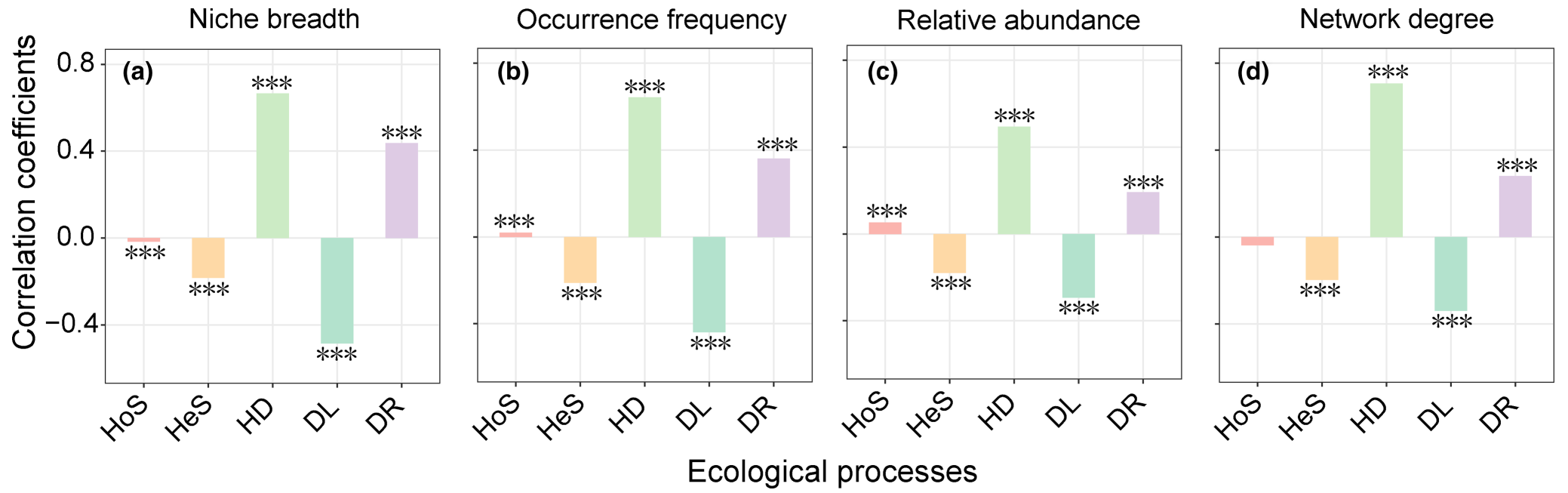
本文系统探究了微生物子群落构建机制与相应微生物分类生态属性之间的关系(图5和图S8)。生态位宽度、出现频率、相对多度和网络度等生态属性与微生物子群落构建机制之间表现出高度一致的关系(图5和图S8)。具体来说,微生物的出现频率、相对多度、网络度和生态位宽度与均质扩散和漂变的贡献呈显著正相关,而与异质选择和扩散限制的贡献呈负相关(图5和图S8)。然而,同质选择的贡献与生态属性之间只存在一些显著但较弱的相关性(图5和图S8)。我们还单独探究了rRNA操纵子拷贝(原核生物生活策略的间接表征指标)与原核生物子群落构建机制之间的关系,但只观察到非常微弱的相关性(图S8)。针对原核生物和真菌类群分别进行的分析也得到了类似的结果(图5和图S9)。然而,真菌和原核生物之间仍然存在一些差异。特别是,网络度与原核生物和真菌子群落构建机制之间呈几乎相反的相关关系(图S9)。有趣的是,总体关系更类似于真菌而不是原核生物(图S9)。
3.4 土壤微生物功能群的子群落构建机制
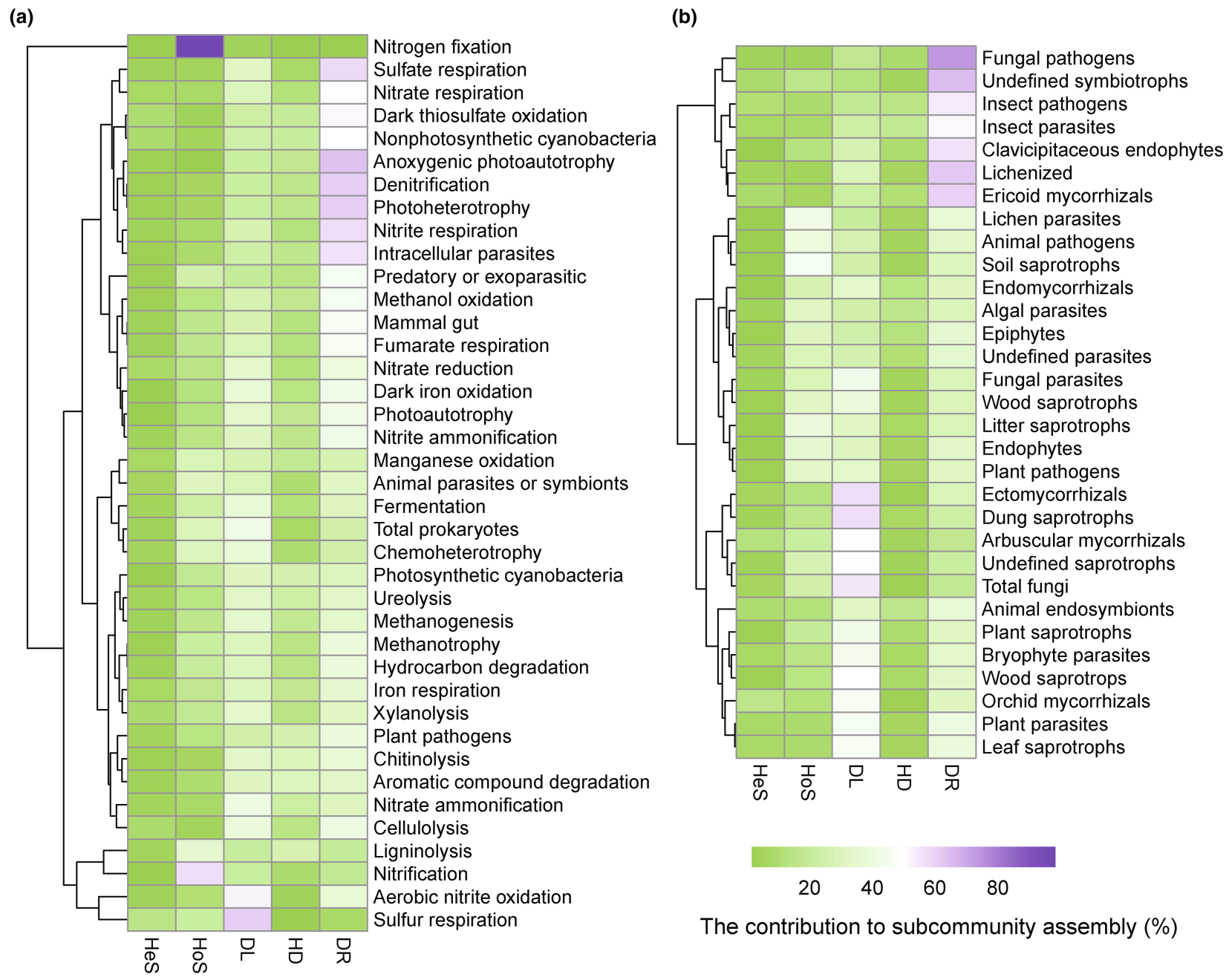
结合FAPROTAX和FUNGuild,我们还观察到不同微生物功能群的子群落构建机制存在显著差异(图6)。大多数原核生物功能群(例如动物寄生虫或共生菌、哺乳动物肠道菌、反硝化菌等)和三分之一真菌功能群(例如昆虫寄生菌、昆虫病原体、动物内共生菌)的子群落构建主要受漂变控制(图6)。扩散限制主导着约三分之一的原核生物功能群(例如硝化菌、纤维素分解菌、木聚糖分解菌)和一半的真菌功能群(例如丛枝菌根、内生菌根、外生菌根)的群落构建(图6)。一些原核生物(例如固氮菌、硝化细菌、与人类有关的微生物)和真菌(例如动物病原体、内生菌、土壤腐生菌)功能群的群落构建主要受同质选择影响(图6)。值得注意的是,同质选择对固氮菌子群落构建的贡献达到了97%(图6)。
4. 讨论
4.1 土壤原核生物和真菌群落的构建
扩散限制主导了云南地区土壤微生物群落的构建(图2a、2b和S2)。土壤微生物群落构建中扩散限制的主导作用越来越受到关注,在山地地区表现得尤为突出。云南省位于典型的山地地区,因此,其复杂的地理结构可以明显限制微生物物种的扩散。这应该是土壤微生物群落构建中扩散限制主导作用的主要原因。这一发现有两个重要启示:首先,最近的研究表明,人类活动可以大大减弱微生物的扩散限制;因此,加剧的人类活动可能会增加该地区有害微生物入侵造成的生态风险;此外,扩散限制可能导致玉米地中一些有益微生物的缺失,因而接种有益微生物或微生物组对改善生态系统健康具有较大的应用潜力。
我们还发现同质选择在土壤微生物群落构建中起重要作用(图2a和2b)。这一发现得到了几项最近研究的支持,并可以解释如下。首先,农田通常经历频繁施肥和干扰,类似的长期管理模式也可以使土壤环境同质化,增加同质选择的影响。这一解释也得到了该地区森林土壤真菌群落构建中同质选择影响较弱的支持(图S2)。此外,一些研究发现,玉米根分泌大量化学物质可以显著影响根际土壤微生物群落,进一步增加同质选择对微生物群落构建的影响。长期的同质选择也会导致土传病原菌的富集。因此,在该研究区域进行轮作对增加土壤微生物多样性和减少土传病原菌是有益的。
尽管土壤真菌和原核生物群落的构建机制表现出大致的相似性,但仍然存在明显差异。特别是,土壤原核生物群落的构建较少受到扩散限制的影响(图2a和2b)。已经有几项研究报道了土壤真菌和原核生物群落不同的构建机制,这可以通过它们固有差异来解释。首先,微生物的扩散通常是以被动的方式发生的,并且在很大程度上取决于每个微生物类群的扩散概率。在我们的研究区域内,土壤原核生物的多度比真菌高两个数量级,导致相比真菌,原核生物具有更高的被动扩散机会。其次,原核生物通常具有较小的细胞大小,这也有助于它们增强的扩散能力。最后,微生物群落构建机制的差异也可能归因于原核生物和真菌与环境因子之间的不同关系。这些发现部分阐明了本研究中土壤微生物亚群构建机制的变异。
4.2 环境因子与微生物群落构建的关系
本研究发现土壤pH是影响原核生物群落构建的主要环境因子(图2c)。这与pH是形成细菌群落的主要环境因子的共识一致,并可以通过大多数细菌相对较窄的最适pH范围来解释。因此,土壤中pH差异的增加可以增强异质选择对土壤原核生物群落构建的影响,并减弱同质选择的影响。有趣的是,本研究所观察到的扩散限制对原核生物群落构建的贡献与土壤pH存在显著的正相关(图2c)。这可能是由pH和地理距离之间的共线性引起的。土壤pH差异进一步限制了微生物的定殖,从而也对观察到的正相关做出了解析。真菌群落构建机制主要与海拔、年均温和年降水呈现出显著但较弱的相关性(图2d)。这可能是因为年均温和年降水等许多环境因素随着地形和海拔的变化而间接与扩散限制的影响相关联。此外,年均温和年降水等气候因子在塑造真菌群落方面发挥着关键作用。这可能是它们与选择对真菌群落构建的贡献之间显著相关的主要原因。实际上,在许多研究中观察到了降水和温度对土壤真菌群落的显著影响。正如上文所述,微生物群落构建机制的驱动机制的差异也部分解释了它们在本研究中不同的亚群构建机制的变化。
4.3 微生物亚群构建机制与生态属性的关系
我们观察到微生物亚群构建机制之间存在着高度异质性,它们与微生物分类单元的生态位宽度和发生频率之间呈现出强烈的相关性(图5a和5b)。具体来说,生态位宽度和发生频率较高的微生物分类单元的亚群构建受到异质选择和扩散限制的控制较少,但更受到均质扩散和漂变的影响(图5a和5b)。这些发现与我们的第一和第二个假设是一致的。微生物分类单元的生态位宽度和发生频率是密切相关的。因此,生态位宽度和发生频率与子群落构建机制呈高度一致的关系。生态位宽度较宽的微生物分类单元通常显示出较高的代谢可塑性,因此受环境选择的影响较小,更多地受到随机过程的控制。相比之下,生态位较窄的微生物分类单元通常具有严格的生长条件要求和低的发生频率,可能伴随着特定的代谢需求,因此更多地受到环境选择而非随机过程的影响。异质选择的贡献与生态位宽度之间存在显著的负相关(图5),表明生态位宽度较窄的微生物更容易受到异质选择的影响。此外,生态位宽度较宽且发生频率较高的微生物分类单元可能具有更高的被动扩散概率。这解释了为何生态位宽度较宽、发生频率较高的微生物分类单元受到扩散限制的影响较小,但更容易受到均质扩散的影响(图5a和5b)。由于iCAMP中定义的漂变包含了太多潜在的生态过程,未来的研究迫切需要进一步调查其与生态属性的关系。这些结果突显了生态位宽度在决定微生物亚群构建机制方面的重要性,并为微生物组学研究提供了新的方向。
有趣的是,我们观察到微生物分类单元的相对多度与异质选择和扩散限制对微生物亚群构建的贡献之间存在显著的负相关(图5c)。在区域尺度上,相对多度较高的微生物通常更适应整体环境,并且对研究区域的环境变化更具抵抗力。这是对微生物分类单元的相对多度与异质选择贡献之间负相关的合理解释。此外,正如上文所述,与稀有微生物分类单元相比,丰富的微生物分类单元具有更高的被动扩散概率。因此,相对多度较高的微生物分类单元的构建受到扩散限制的影响较小,但更受到均质扩散的控制。事实上,一些微生物稀有种可能由于其主动的移动能力或寄主而具有较强的扩散能力。然而,由于大部分微生物主要通过被动扩散传播,这种解释至少在大多数情况下仍然得到支持。这些结果直接将微生物相对多度与微生物子群落构建机制联系起来,为不同稀有程度的微生物子群落构建机制提供了关键的见解。
本研究中的另一个有趣发现是,具有更高网络关联度的微生物分类单元的构建受到异质选择和扩散限制的影响较小,但更受到均质扩散的驱动(图5d)。一个可能的解释是,具有更强相互作用的微生物分类单元可以形成合作聚集体,因此对环境变化更具抵抗力,并且不太受异质选择的影响。复杂的相互作用模式可以提高微生物对环境干扰的抵抗力。因此,网络度较高的微生物更具竞争力和多度,因此具有更高的扩散概率,并且受到扩散限制的影响较小。然而,当分别分析原核生物和真菌时,我们发现总体关系更类似于真菌而不是原核生物(图S9d和S9h)。一个可能的解释应该是网络度对真菌和原核生物的不同生态意义。由于细胞大小较小,土壤原核生物通常具有比土壤颗粒更小的相互作用半径。因此,网络分析可能无法描述原核生物分类单元之间的相互作用。相反,菌丝的存在使得真菌-真菌和真菌-原核生物分类单元之间的相互作用可以被网络度所反映。这些发现表明微生物子群落构建机制也部分取决于微生物分类单元之间的相互作用,并突显了不同微生物分类单元网络属性的潜在不同生态意义。
4.4 不同微生物功能群的子群落构建机制
根据我们的假设,微生物群落的构建机制在不同功能群之间表现出显著差异,这可能是由多方面因素引起的(图6)。首先,功能群的生态特征(例如细胞大小和基础代谢特征)的差异可能是不同子群落构建机制差异的主要原因。例如,许多原核和真菌功能群(如化学异养生物、腐生菌和外生菌根真菌)的子群落构建机制与总原核和真菌的机制显示出高度相似性(见图6)。此外,正如上文所述,与真菌相比,原核生物普遍较小的细胞大小和更高的多度也使得原核功能群受到的扩散限制较少。其次,许多功能微生物与宿主有着密切的关系,因而一些微生物功能群的子群落构建也会受到宿主的影响。因此,植物相关的功能群(如外生菌根、丛枝菌根和木质腐生菌)主要受到扩散限制的影响,而动物相关的真菌功能群(如昆虫寄生真菌、昆虫病原真菌和动物内共生微生物)主要受漂变的影响(图6)。有趣的是,固氮菌主要受同质选择的影响(97%;图6),这可以由它们与玉米的共生共生关系所解释。第三,一些微生物功能群的子群落构建也可以受其特定功能特征的控制。例如,已有研究表明,施肥是影响生物固氮的关键因素。因此,同质选择对固氮菌群的显著影响也可能是目前广泛采用的施肥所造成的。硝化菌子群落构建机制也可以用类似的方式来解释。总之,不同功能群展示了多样的微生物群落构建机制,归因于它们的基本特征、宿主和特定功能特性。这些发现对耕地管理具有重要意义。例如,我们应该通过接种解除植物促生菌的扩散限制,同时限制有害菌的扩散。
5. 结论
本研究表明,扩散限制和同质选择在云南土壤微生物群落构建中起着主导作用。气候和pH分别是预测真菌和原核生物群落构建机制的主要驱动因素。不同微生物分类群的子群落构建机制也有所不同,并且它们与生态位宽度、出现频率、平均相对多度和网络关联度之间存在显著相关性。异质选择和扩散限制的相对贡献与微生物分类群的出现频率、平均相对多度、网络度和生态位宽度呈负相关。相反,均质扩散和漂变的相对贡献与这些生态属性呈正相关。本研究还观察到不同微生物功能群的子群落构建机制存在显著差异,这主要可以通过它们的生理特征以及与宿主的关系来解释。总而言之,本研究揭示了土壤微生物子群落的构建机制,以及它们与特定分类群的生态属性和功能特征之间的联系,为探索土壤微生物多样性维持机制提供了新的方向。此外,本研究开发的微生物功能子群落构建机制分析流程也可为未来微生物组研究提供有力工具。
-
点赞 (0人)
- 收藏 (0人)

