
iMeta | 南方医科大学谢黎炜、马颖联合发现新生儿肠道微生物网络结构变化规律
- 看不见的线
- 5225
- 2024-06-07 10:14:06
- 文章来源:iMeta
新生儿湿疹肠道菌群网络结构图谱

iMeta主页:http://www.imeta.science
研究论文
●原文链接DOI: https://doi.org/10.1002/imt2.90
●2023年2月16日,南方医科大学谢黎炜、马颖团队在iMeta在线发表了题为“Microbial network signatures of early colonizers in infants with eczema”的文章。
●本研究结合图论与网络算法系统探索了中国婴儿生命早期的肠道菌群变化,联合分析结果表明不同民族的健康婴儿在生命初期的肠道菌群网络形态发展规律相似;并且健康婴儿的肠道菌群网络形态比患有湿疹的婴儿更稳定和复杂。此外,健康的婴儿肠道菌群丰富度在生命早期逐步增高,而母亲在孕晚期血清甘油三酯(TG)的高水平可能是出现新生儿湿疹的危险因素。
●第一作者:黄柳菁、盘桂花
●通讯作者:谢黎炜 (xielw@gdim.cn)、马颖(mayingwuzhuoyi@126.com)
●合作作者:冯译菲、范紫晶、马凯、王润鑫、王广贤、黄广业、黄思霞、侯语汇、韩木兰
●主要单位:南方医科大学珠江医院妇产医学中心;广东省科学院微生物研究所,华南应用微生物国家重点实验室;新乡医学院公共卫生学院;江苏新申奥生物科技有限公司
亮 点

●新生儿胎便菌群已具备一定的网络拓扑学复杂度,随着肠道菌群的成熟,菌群网络复杂度呈增加趋势
●生命早期健康婴儿的肠道菌群网络结构比患有湿疹的婴儿更复杂和更稳定
●妊娠晚期甘油三脂水平升高可能是后代生命早期出现湿疹的危险因素
摘 要
既往研究表明肠道菌群与新生儿湿疹存在关联关系,且患有湿疹的婴儿肠道菌群出现失衡与紊乱。尽管婴儿肠道菌群组成已相对明了,但菌群内部构成的网络结构及其与宿主生理和病理表型的关系亟待研究。为了研究生命早期100天的肠道菌群网络结构变化,我们在广州开展了一项队列研究,一共纳入了34位足月阴道顺产的婴儿,并且采集了胎粪以及出生后3个月共计四个时间点的粪便样本,并进行16S rRNA扩增子测序和数据分析。同时,我们额外纳入了两项队列研究的健康婴儿肠道菌群样本来支持本研究的结果。本研究结合图论与网络算法系统探索了中国婴儿生命早期的肠道菌群变化,此外,联合分析结果表明不同民族的健康婴儿在生命初期的肠道菌群网络形态发展规律相似;并且健康婴儿的肠道菌群网络形态比患有湿疹的婴儿更稳定和复杂。此外,健康的婴儿肠道菌群丰富度在生命早期逐步增高,而母亲在孕晚期血清甘油三酯(TG)的高水平可能是出现新生儿湿疹的危险因素。
视频解读
Bilibili:https://www.bilibili.com/video/BV1Rj411V7xF/
Youtube:https://youtu.be/dCsYZ_PW8jA
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
湿疹(也称为特应性皮炎,AD)是一种免疫相关的炎症性皮肤病,经常在生命早期发展并持续到成年期。根据全球疾病负担调查,这种疾病影响了至少 15% 的儿童,并且其患病率逐年上升。该病的症状与年龄、疾病分期、种族和地理位置密切相关。在婴儿中,湿疹通常会影响头部、面部和四肢,导致皮肤干燥、开裂和鳞屑。
免疫印记可以追溯到胎儿时期,并与母体微生物群及其代谢物相关,例如短链脂肪酸 (SCFA),已被证明在免疫调节中具有重要作用。微生物组成与人类健康密切相关,新生儿可通过接触丰富的微生物并获益。先锋物种的初始定殖始于出生,随后微生物多样性和群落迅速增加。在生命的前 100 天,婴儿主要从母乳或配方奶中获取营养。一项大规模的荟萃分析发现,如果在出生后的前三个月进行纯母乳喂养,具有AD家族史的儿童患AD的可能性较小。由于微生物多样性与人类健康之间存在的密切关系,了解微生物群落的形成和维持对于靶向整个微生物群落的干预是十分重要的。
微生物群对宿主生理学影响的研究主要包括四个方面,即屏障功能、免疫调节、定植抵抗和发育。部分微生物可产生有益的代谢物并发挥功能,这些共生微生物的稳定性被认为是宿主健康和机体维持稳态的关键。因此,肠道微生物群的急剧变化往往与疾病的发生和发展密切相关。此外,在肠道生态系统中存在诸多细菌相互作用,通过在生态位内代谢物、能量、信号和其他物质的交换,各种微生物共同形成一个复杂的生态系统。为了剖析这个生态系统,网络算法和图论已经被用来揭示微生物群落内的竞争、共生和互动情况。
尽管临床研究已经报道了肠道微生物群的改变与儿童湿疹患病率之间的关联,使人们对儿童湿疹的微生物群失调有了深刻的了解,但仍然缺乏肠道菌群网络结构在生命早期如何发生变化以及其与宿主表型关系的研究和探索。采集婴儿期到幼儿期的粪便样本进行纵向分析,可以动态的认识生命早期微生态的变化。尤其是生命早期100 天肠道菌群的组成和变化对免疫印记至关重要,而已发表的研究并未将此时间段作为研究重点。此外,母亲孕前体重指数 (BMI) 对后代湿疹发展的影响仍存在争议。因此,有必要使用网络和图论等方法深入探索肠道微生物组与湿疹风险之间的关系。
为了更好地了解婴儿出生后前 100 天的肠道微生物网络,我们在南方医科大学珠江医院妇产医学中心招募,并采集了婴儿出生以及出生后1、2 和 3 个月龄的粪便样本,进行 16S 扩增子测序和数据分析,系统的探索了与湿疹相关的潜在微生物群因素。
结 果
母婴人口学特征及临床检查结果
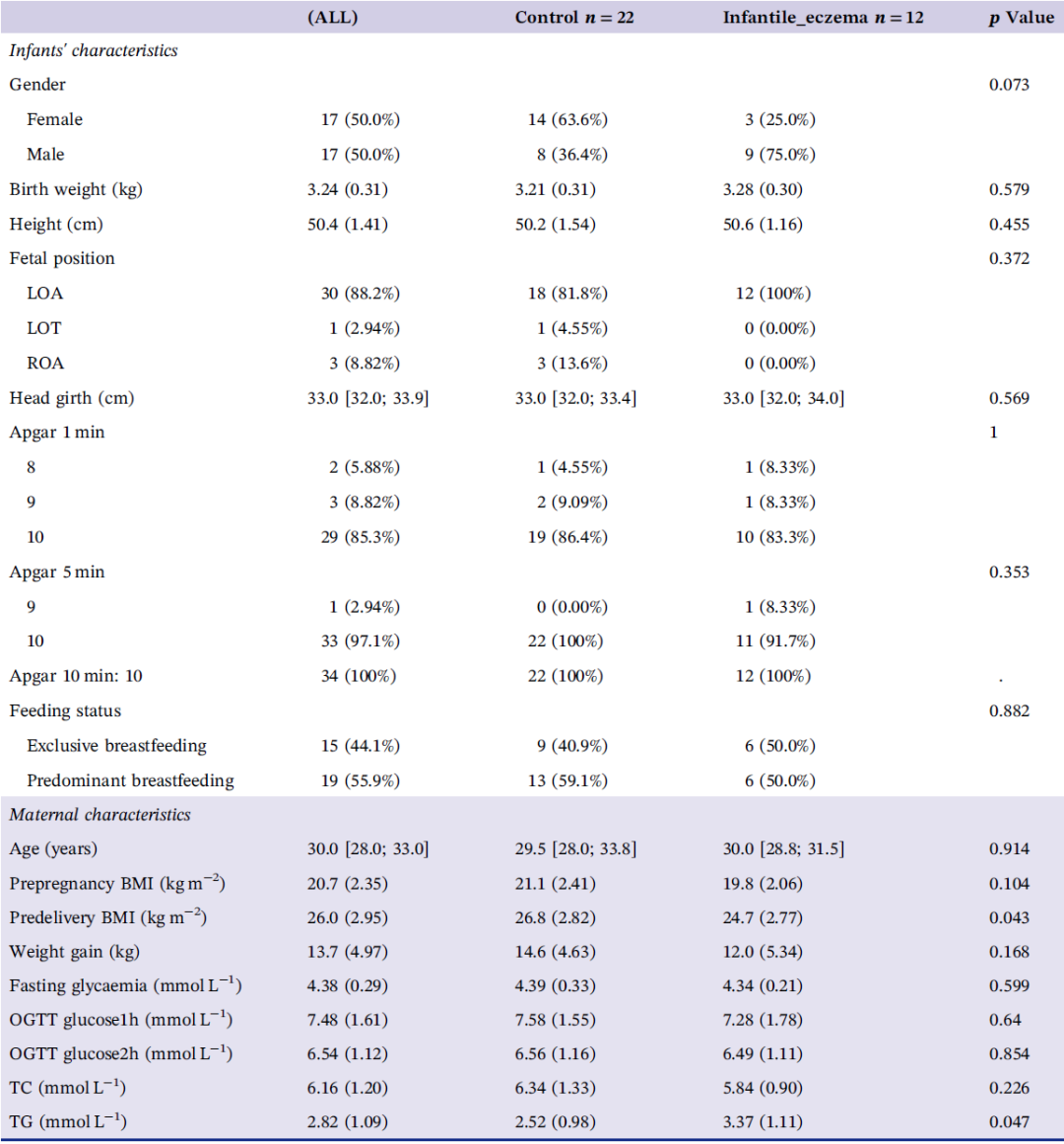
在妇产医学中心出生的新生儿,如果常规检查无异常,则不进行任何血清学或影像学检查。婴儿性别、出生体重、身长 、头围、胎儿分娩方式和 Apgar 评分在对照组(Healthy Control)和病例组(Infantile eczema Group)之间的差异没有统计学意义。两组婴儿都是纯母乳喂养或主要母乳喂养,并且在采样期间孕妇和胎儿没有接受抗生素治疗。入选的34位妈妈均居住在广州,孕期均定期进行产检。所有产妇入院后均接受了全面的体格检查以备分娩,包括血常规检查、凝血功能、肝肾功能、口服葡萄糖耐量试验(OGTT)、感染性疾病、血型、甲状腺功能和糖化血红蛋白水平等。这些定期检查的临床指标,包括孕前体重指数(BMI)和孕期体重增加情况,在对照组和病例组之间没有发现显著差异。然而,病例组婴儿的母亲分娩前BMI明显低于对照组(表1;表S1)。既往研究表明,孕前 BMI 和孕期体重增加均与围产期并发症和不良妊娠结局的发生有关。在本研究中,虽然母亲孕前 BMI 在两组间不存在显著差异,但两组婴儿的母亲在分娩前 BMI 存在显着差异。此外,病例组婴儿的母亲在分娩前的血肌酐和甘油三酯 (TG) 水平显著升高。两组母亲的肌酐均在正常范围内(表 S1)。然而,因为怀孕期间激素的变化会导致甘油三酯的生理性升高,两组母亲的TG均高于正常范围。但有研究提示,妊娠晚期母亲血清中甘油三酯水平高可能与妊娠不良结局事件有关。
表1. 受试者婴儿及母亲的人口学特征及临床检查结果
健康婴儿和湿疹婴儿的肠道微生物多样性不同
对照组和病例组门级微生物群的堆积条形图如图 1B 所示。婴儿肠道中的主要先锋菌在门水平包括变形菌门、厚壁菌门、放线菌门和拟杆菌门。其中,以蓝色显示的变形菌门和厚壁菌门在婴儿胎粪样本中为优势菌,而变形菌和放线菌门在出生后第 1、2 和 3 个月的粪便样本中占主要构成。Alpha 多样性被用于描述肠道生态系统的微生物丰富度和均匀度。如图 1C 所示,与胎粪相比,在婴儿第 1、2 和 3 月龄粪便样本中,对照组肠道菌群的Inverse Simpson (InvSimpson) 指数显著升高。与对照组相比,病例组1、2和3月肠道菌群样本的 InvSimpson 指数较胎粪样本有增加趋势,但未达到统计学差异(图 1C)。基于主坐标分析(PCoA)的Beta多样性结果表明,婴儿第1、2和3个月的肠道微生物群结构与胎粪之间的差异具有统计学意义(图 1D)。Beta多样性结果与细菌相对丰度的堆叠条形图中观察到的结果相互呼应(图 1B),在图 1C中我们可以观察到在第 1、2 和 3 个月收集的粪便样本和胎便样本之间的明显差异。为了进一步验证在不同时间点收集的样本中微生物结构的组间变异性,对不同时间点样本分别进行了降维分析。如结果所示,两组婴儿的胎粪样本(图 1E,p= 0.1)、产后第 1 个月的粪便样本(图 1F,p= 0.96)和产后第 2 个月(图 1G,p为 0.084)。在 PCoA 轴 2 的组间变异性逐渐变大,说明健康婴儿和出现湿疹的婴儿的肠道微生物群结构随着发展和成熟过程中逐步出现差异。有趣的是,在产后第 3 个月观察到 PCoA 轴 2 的差异具有统计学意义(图 1H,p= 0.011)。这些结果表明,肠道微生物群的组成在婴儿期逐渐发生变化,对照组婴儿的肠道菌群丰富度增加更为显著。
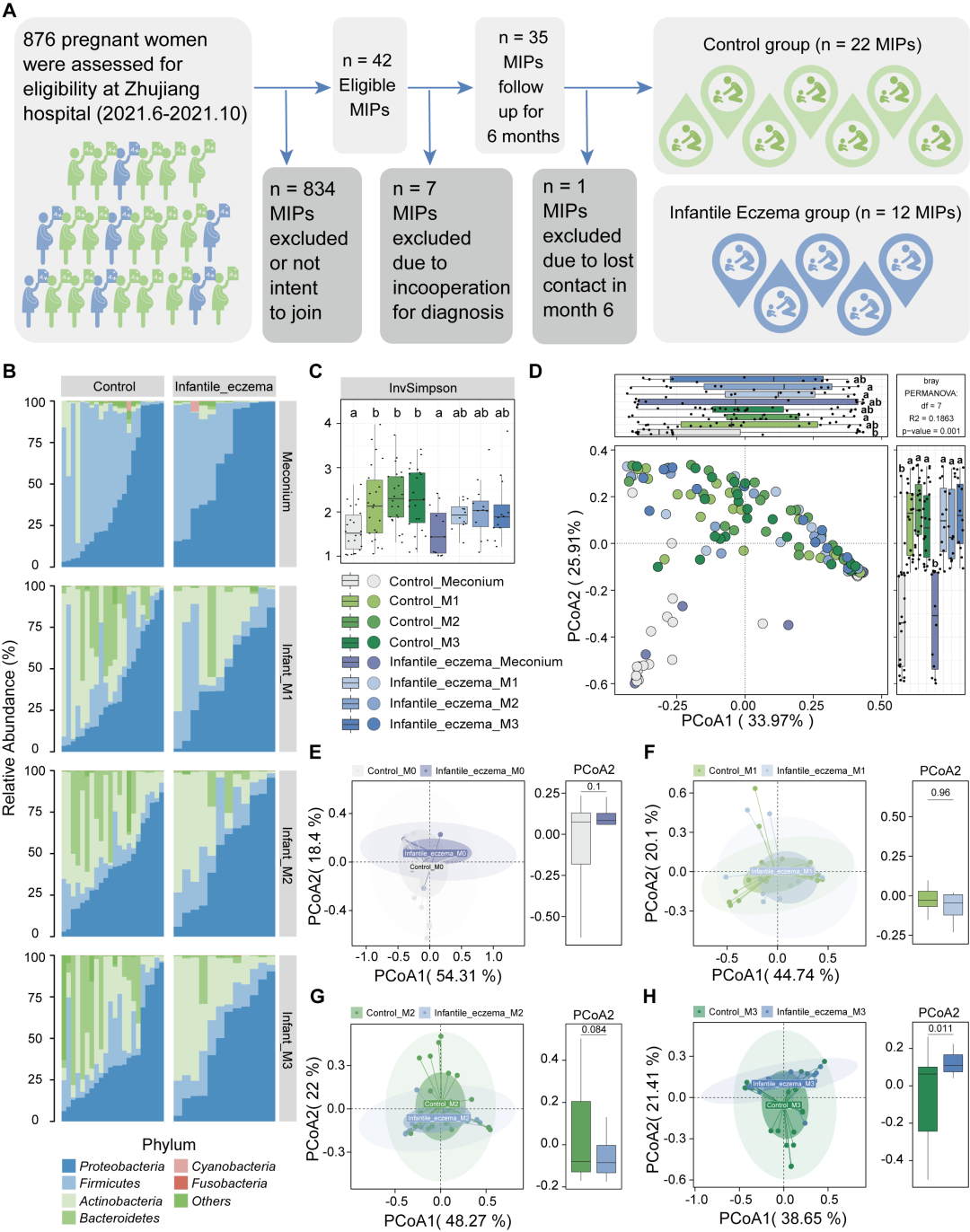
图 1. 生命早期婴儿肠道菌群结构特点
(A)研究流程图;(B)菌群堆积条形图;(C)基于Inverse Simpson指数的Alpha多样性箱线图;(D)基于PCoA (Principal Coordinate Analysis)的β多样性降维分析结果;(E)胎便样本的β多样性;(F)1月龄样本的β多样性;(G)2月龄样本的β多样性;(H)3月龄样本的β多样性。
健康婴儿在生命早期微生物网络具有相似的节律性
生态网络的复杂性对生态系统稳定性具有重要作用,而我们对新生儿肠道微生物群网络形态的了解不足。因此,我们利用受试婴儿生命早期共计4个时间点的粪便样本开展生命早期肠道微生物群网络的结构研究。我们基于每组样本的扩增子序列变异 (Amplicon sequence variant, ASV) 构建了时间序列分子生态网络 (MEN)(图 2A)。为了更好的说明肠道菌群在生命早期的网络结构变化,我们整合了Roswall 等人的瑞典队列和Christensen 等人的丹麦队列中随机选择的 16 名健康婴儿,获得了在不同时间点收集的肠道菌群样本,并构建 MEN(图 S1A、B),我们可以观察到生命早期肠道微生物群中存在的交互网络。网络拓扑学指标用于展示视觉映射,包括簇数(Number of clusters, No. Clusters)、边数(Number of edges, Num. Edges)、正相关边数(Number of positive edges, Num. Pos. Edges)、负相关边数(Number of negative edges, Num. Neg. Edges)、直径(Diameter)、平均路径(Average path length), 中介中心性(Centralization betweenness),其中几个网络图论指数在胎便样本中大于零,表明健康新生儿肠道中已经存在一定程度的微生物群网络(图2B)。
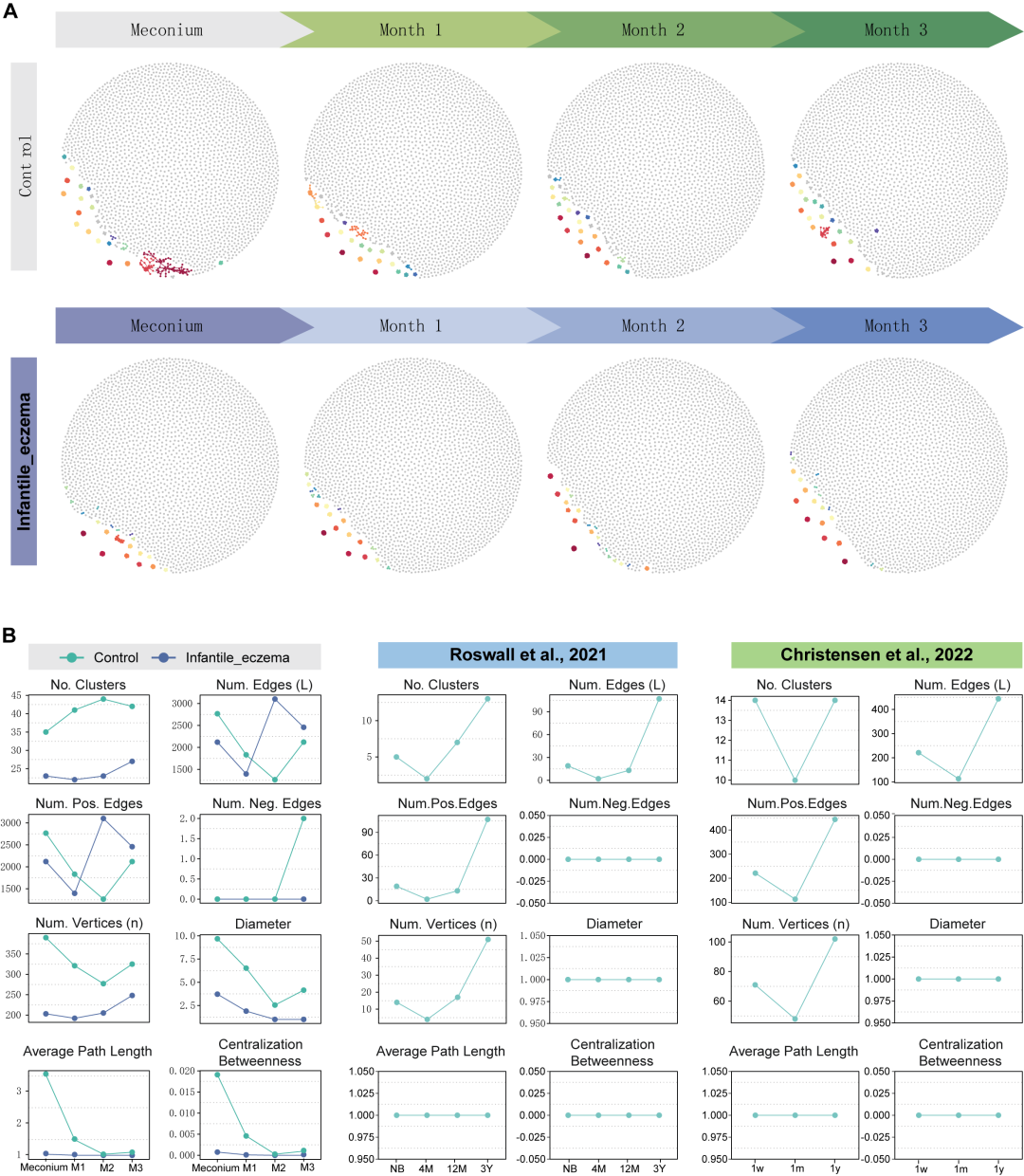
图2. 生命早期肠道菌群网络结构变化
(A)时间序列分子生态网络 (MEN);(B)网络拓扑学指标,折线图分别反映了本研究中的健康婴儿(绿色)和患有湿疹的婴儿(蓝色),以及两个独立研究的健康婴儿(绿色)肠道网络结构变化。
降低生产成本并保护环境
从结果中,我们可以观察到健康婴儿早期先锋菌网络拓扑结构的复杂性已具备一定水平,例如簇数、边数、正相关边数、顶点数、平均度(Average degree, Average K)、相对模块化(Relative modularity, RM)、集中度(Centralization degree)(图 2B;图 S1C)。从绿色的折线图中,我们可以观察到这些指标从第二个采样时间点开始逐渐增加,提示网络随着时间的推移变得复杂和成熟。在 Roswall 等人和 Christensen 等人的研究提取的独立队列数据分析结果中,这些网络拓扑学指标仍然为零,例如直径、平均路径和中介中心性;但是当前队列中健康婴儿的结果可以观察到一定变化趋势。我们推测这是由于 16S rRNA 数据的测序质量差异所致。在我们的研究中,由于采用了标准严格的采样流程、测序流程以及主流测序仪,我们获得了高质量的测序数据。相反,对独立队列的16S rRNA数据进行质量控制过程中过滤了不合格的序列,导致下游分析的ASV数量有限。为了进一步探索独立队列的基本微生物特征,我们分别绘制了菌群堆积条形图并且进行了alpha 多样性和 beta 多样性分析(图 S2)。从图中我们可以得出结论,虽然健康婴儿的肠道微生物特征在三个数据集中有所不同,但网络拓扑指数在新生儿胎便样本中处于较高水平,并且网络复杂性随着肠道微生物群的成熟而逐渐增加。值得注意的是,三批时序数据的keystone节点数指标均为零,反映出虽然生命早期肠道内已经形成了稀疏的网络结构,但目前还没有满足算法界定的核心先锋菌。此外,在本研究婴儿3月龄采集的肠道菌群中观察到负相关边数的出现,反映了先锋菌之间的竞争。总之,这些数据表明网络拓扑指数在胎便样本中较高,并且随着肠道菌群的成熟可以观察到网络复杂性增加的趋势。上述结果表明,健康婴儿的肠道微生物网络存在节律性变化,这可能对生命早期的健康发展至关重要。
健康婴儿的肠道微生物群网络更密集
人体肠道微生物群存在内部相互作用并且微生物群的网络稳定性对宿主健康至关重要,但目前对网络稳态仍知之甚少。婴儿时期的肠道微生物群一直是人们关注的焦点,这是肠道微生物群和其他生理和生物学过程发展的一个重要时期。因此,探索生命早期100天的微生物群的互作并识别其变化规律有利于加深我们对新生儿湿疹的认识。在本研究中,相同时间点对照组婴儿肠道菌群的簇数高于病例组婴儿(图 2B)。另外,对照组婴儿肠道菌群簇数自出生起开始增加,第3个月略有减少,但仍高于第1个月和胎便样本。而病例组婴儿肠道菌群簇数在出生后减少,然后在第3个月显著增加。对照组健康婴儿肠道菌群网络图中边数、正相关边数和顶点的数量先减少后增加,这个动态模式也在两个独立数据集中得到验证。而在病例组中边数和正相关边数在第三个月明显下降,而顶点数则在生命早期趋于平稳。由于在独立数据集中获得的顶点数量较少,MEN直径在这种映射模式下没有改变。另一个有趣的结果是,对照组婴儿3月龄时,肠道菌群可观察到负相关的边,反映了微生物群落中竞争拮抗簇的出现,但在病例组中没有观察到该现象。不仅如此,代表病例组的蓝色折线在多个网络拓扑指数中的变化均出现异常。例如,边数、正相关边数、连接性(Connectance , Edge density)、平均度、接近中心性(Centralization closeness)、中心度(Centralization degree)均在第二个月出现高峰,并在第三个月骤降,这种骤变可能与婴儿湿疹的发病有关(图 2B;图 S1C)。总的来说,我们发现湿疹婴儿的网络特征节律缺失,而健康婴儿的肠道微生物群网络更加密集。
婴儿肠道菌群的建立
菌株水平研究发现,母婴之间菌株的垂直传播是婴儿肠道先锋菌的主要来源。鉴于 16S 扩增子测序在物种水平鉴定准确性的不足,本研究中,我们分析比较了不同时间点两组样本间菌科的差异,但没有观察到任何具有统计学意义的差异菌。该结果可能是由于本项研究招募的婴儿均为阴道分娩,并且在出生后的前 3 个月内采取纯母乳喂养或者母乳喂养为主仅偶尔补充配方奶粉。三元图被用于展示婴儿肠道中先锋菌的发展转化过程(图 3A、B)。在对照组和病例组的胎便、产后1月和产后3月肠道菌群样本中,肠杆菌科和双歧杆菌科都是生命早期的优势细菌(表 S2 和 S3)。从三元图中灰色线条内部区域可见到对照组和病例组分别只有 2 个和 5 个优势菌科从出生持续到第 3 个月,反映了生命早期肠道微生物群的剧烈变化(图 3A、B)。乳杆菌科在胎便样本中占比更高(图 3A,B),开始接受哺乳后采集的肠道菌群中乳杆菌科的占比不及 1%(图 3C)。肠杆菌科是一类兼性厌氧菌,是人类和动物的共生菌之一。另一方面,双歧杆菌科和乳杆菌科在生命早期展现出相反的变化趋势。长双歧杆菌、动物双歧杆菌和鼠李糖乳杆菌等均是在这两个益生菌科下的有益菌,在既往大量研究中被报导能够改善机体健康。在本研究中,我们观察到婴儿粪便中乳杆菌科的相对丰度在母乳喂养开始后显著下降(图 3C)。相比之下,双歧杆菌科在哺乳开始后在肠道中更为丰富(图 3D)。从三元图中,我们还可以得出结论,健康婴儿和患有湿疹的婴儿肠道菌群的发育模式存在很大差异。

图3. 婴儿肠道菌群的建立
(A)健康婴儿胎便、出生1月和2月肠道菌群三元图;(B)患有湿疹的婴儿胎便、出生1月和2月肠道菌群三元图;(C)乳杆菌科相对丰度箱线图;(D)双歧杆菌科相对丰度箱线图。
母亲妊娠晚期甘油三酯水平升高可能是湿疹的危险因素
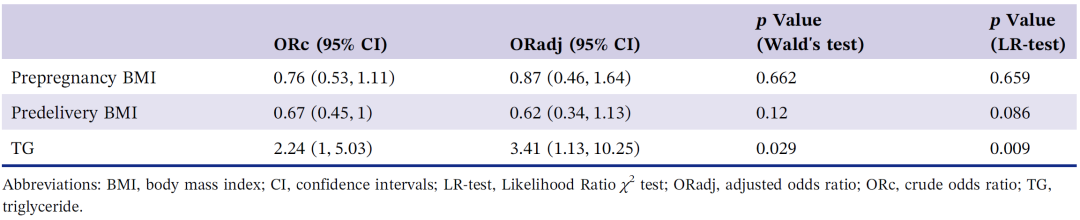
根据变量特征采用独立t检验、Mann-Whitney检验、卡方检验和Fisher精确检验对两组婴儿基本特征、母体人口统计学和临床检查结果进行单变量比较。为了确定与湿疹存在独立相关的变量,选择单变量分析中 p< 0.05 的变量作为多元逻辑回归的潜在自变量。产妇孕前BMI统计结果虽然未达到0.05的统计阈值,但对产前BMI有直接影响。因此,将孕前BMI一并纳入到多元逻辑回归分析。统计结果如表2所示,湿疹的发生与母亲孕晚期 TG 水平呈正相关(校正后的比值比 = 3.41 (95% CI: 1.13-10.25),p= 0.029)。因此,母亲妊娠晚期甘油三酯水平高可能是湿疹的危险因素。
表2. 多元逻辑回归
讨 论
我们以间隔一月的采样频率收集了婴儿出生后4个时间点的粪便样本,并完成了定期随访以获取每对母婴的详细信息。联合另外两项独立纵向调查的 16S rRNA扩增子测序数据,我们发现新生儿胎便菌群已具备一定的网络拓扑学复杂度,随着肠道菌群的发育,菌群网络复杂度呈增加趋势。此外,基于我们采集的数据发现健康婴儿的肠道微生物群网络比患有湿疹的婴儿更密集。我们的研究结果还表明,肠道微生物群的生态演替在婴儿期是一个动态过程,健康的婴儿在生命早期100天的肠道微生物丰富度增加明显,母亲妊娠晚期血清中甘油三酯水平升高可能是新生儿湿疹的危险因素。
越来越多的证据表明,肠道微生物群对于免疫系统的发育以及新生儿、婴儿、儿童和成人时期湿疹的预防至关重要。其中,一些横断面研究获得了 6 个月以下婴儿的粪便样本,而其他研究则进行了纵向研究。一项纵向研究通过对 5 周龄至 6-11 岁儿童粪便微生物群进行连续采样,数据表明微生物多样性和组成的改变要早于过敏的出现。此外,肠道菌群自出生时建立,随后微生物多样性和群落迅速增加。这些研究强调了生命早期微生物演替和成熟的重要性,但是研究中纳入出生后 100 天婴儿粪便的设计并不多。只有一项研究覆盖了来自产后 5、13、21 和 31 周以及6 至 11 岁儿童(婴儿)的样本;然而,该研究采样间隔大,而且没有采集胎便样本。在我们的研究中,以一个月的采样间隔,在四个时间点收集婴儿粪便样本,使我们能够探索生命早期的微生物特征和成熟。此外,最近发表的一项研究表明,没有证据表明胎便菌群组成与 4 岁以下儿童的特应性皮炎有关。在本研究中,胎便菌群结构在组间未观察到明显差异,但从第 2 个月的肠道菌群中可以观察到差异(图 1G,p = 0.084),并在第 3 个月达到统计显著性(图 1H,p = 0.011)。总的来说,在控制了包括出生方式、母乳喂养状态和抗生素使用情况这些混杂因素后,我们发现患有湿疹的婴儿在出生后的头 100 天内的 alpha 和 beta 多样性发展情况与健康婴儿相比存在异常。
在漫长的历史中,图论作为数学的一个分支,以图为研究对象进行节点和边关系的数学推理。随着社交网络分析和网络科学的发展,网络研究得到越来越多的关注,图论也开始应用于这些领域。然而,新生儿微生物组研究大多缺乏对菌群网络特性的进一步描述。共现网络是最常用的网络类型之一,它可以直观地显示不同微生物类群之间的正相关和负相关关系。虽然共现网络可以显示不同分微生物类群之间的复杂关系,但我们不能忽视一个事实,即不存在一个完全适用的健康微生物组配置。ASV 是微生物的预注释核酸序列,可以更好地应用于构建网络结构以证明生态系统内的相互联系。此外,在基于人群的宏基因组学分析中,作者证明 16S rRNA 基因测序在预测微生物科级别结果要多于宏基因组鸟枪法测序。结合网络拓扑指标与ASV结果能共同体现网络的复杂性和稳定性。为了验证我们的假设,即生命早期肠道微生物网络节律的动态变化与婴儿的健康密切相关,我们基于三个独立数据集的科水平的 ASV 构建了 MEN,并证明了不同国家的健康婴儿的网络拓扑指标共享一致的动态变化;并说明了与患有湿疹的婴儿相比,健康婴儿具有更密集的肠道微生物群网络。
湿疹的原因是多方面的,包括遗传、父母和生活环境等。在这些因素中,婴儿的肠道微生物组对儿童免疫系统的发育具有重要影响。在本研究中,InvSimpson 指数在生命早期的健康婴儿中显著升高,而患有湿疹的婴儿只能观察到升高的趋势但变化没有统计学意义(图 1C)。结果表明 InvSimpson 指数可能是一种保护因素,这与之前的研究一致。InvSimpson 指数是能够反映微生物组丰富度和均匀度的 alpha 多样性指数之一。其中一个可能的原因是两组婴儿的生活环境不同。较早的一项研究发现,父亲受过高等教育的婴儿患湿疹的风险更高(ORadj(95% CI)= 9.93(1.83-53.71),p= 0.008)。研究人员还推测,受教育程度较高的父亲可能对卫生保洁有更高的要求,这表明婴儿在早期因接触不卫生环境而感染传播性疾病的概率较低,但这可能是过敏性疾病的患病率上升的原因。另一项研究调查了城市化微生物群对婴儿、免疫体质和特应性疾病发病的影响,表明城市化引起的婴儿微生物群的变化可能会通过影响免疫系统的发育而增加患哮喘和特应性皮炎的风险。另一方面,越来越多的研究调查了农村生活对婴儿成长的有益影响,强调由于城市化发展,婴儿不能暴露于某些微生物的环境导致了过敏性疾病的全球上升。尽管在本研究中的婴儿均在中国,广州,这个城市化程度高的城市出生和长大,但他们的生活环境可能仍然不同。上述研究发现可以通过接触非城市化环境来提高肠道微生物多样性,这可能是降低过敏性疾病风险的一种方法。在目前的研究中,从α多样性和网络特性的指标,我们可以观察到湿疹婴儿肠道菌群的发育出现了异常。我们推测肠道中早期定植菌的丰富性对于发展复杂的微生物网络可能很重要。据报道,益生菌、益生元和合生元在预防和治疗许多健康问题都表现出一定的潜力,我们推测对于肠道微生态失调的湿疹婴儿适当的补充微生态制剂有望通过增加肠道菌群的丰富度来促进微生物网络的进展。除了可能与环境因素密切相关的InvSimpson指数外,目前的研究发现,母亲孕晚期血清甘油三酯升高可能是湿疹的危险因素(ORadj(95% CI)=3.41(1.13-10.25) , p = 0.029)。虽然两组母亲产前BMI存在显著差异,但多元逻辑回归分析表明,只有孕晚期产妇TG高是湿疹发生的危险因素。既往研究中,有研究发现母亲肥胖和妊娠期体重增加不是特应性疾病的危险因素,同时也有报道称母亲孕前BMI是儿童期湿疹的危险因素之一。其中的机制仍需要进一步研究阐明。
这项研究存在一定的局限性。首先,它不是基于大样本的研究,往后应该进一步扩大队列。季节变化和不同的气候因素对湿疹的发病和严重程度存在一定影响。虽然本研究的婴儿是在中国广州的夏季招募的,当时气温在 30 摄氏度左右,但我们没有对婴儿的成长环境进行调查。此外,湿疹被认为与整个婴幼儿时期的睡眠质量受损有关,这强调了长期跟踪受试婴儿的疾病严重程度、同期出现的症状以及睡眠质量和睡眠持续时间的重要性。最后,如果能够持续每日采样将能够更细致地发现肠道微生物初期演替过程中网络复杂性发展的规律。
结 论
网络分析和图论可用于发现微生物群的形成和构建过程。在本研究中,不同种族的健康婴儿在生命早期具有相似的肠道微生物网络变化,而与健康婴儿相比,患有湿疹的婴儿则表现出更稀疏和波动的网络。通过时间序列数据,我们证明湿疹婴儿的微生物结构在第 3 个月出现明显失调。研究结果还提示母亲妊娠晚期血清甘油三酯水平升高可能与婴儿湿疹的发生有关。
方 法
受试者入组
2021年6月至2021年10月,对中国广州市南方医科大学珠江医院产科收治的所有孕妇进行了健康评估。先前的研究表明,婴儿出生方式会显著影响肠道菌群的结构,在阴道分娩过程中婴儿通过垂直传播途径接触到母亲阴道中的乳杆菌。因此,我们仅招募了阴道分娩的婴儿。此外,世界卫生组织 (WHO) 和联合国国际儿童紧急基金会 (UNICEF) 建议儿童在出生后的第一个小时内开始母乳喂养,并在出生后的前 6 个月内进行纯母乳喂养。因此,本研究仅纳入了具有纯母乳喂养意愿的孕妇。由于个体间存在微生物群的异质性,我们在入组期间设计了严格的标准(如入组排除条件所示),尽管在本市的三甲医院展开研究,4 个月内共入组的队列规模仍然较小。在当前的研究中,我们对 876 名健康孕妇及其婴儿进行了初筛,其中大部分因不符合入组标准或拒绝参加本研究而未被纳入。进入采样流程的母婴共计42对。生命中的早期定殖者对健康至关重要,尤其是对婴儿免疫系统的发育。(新生儿)胎便主要在胎儿时期形成,而胎儿处于相对厌氧的环境中,胎粪样本中兼性厌氧菌和严格厌氧菌的比例明显高于后期采集的婴儿粪便。由于免疫系统的发育可以追溯到胚胎期,我们推测胎便的网络结构可能与免疫系统的发生有关。
因此,分娩后婴儿的胎便样本由医生在医院采集。我们给所有的母亲发放了居家采样指南,介绍了在家中收集产后第 1、2、3 个月婴儿粪便样本的正确方法。我们在产后前3个月提供健康指导并记录母婴的基本信息,例如健康状况和药物使用情况。在婴儿 6 月龄时再次通过问卷收集婴儿的健康状况信息。然而,在采样期间,有 7 对母婴因拒绝继续配合或诊断出疾病而退出研究。所以最终,我们从 35 对母婴处获得了4个时间点的样本,并对这些样本进行了 16S rRNA 基因扩增子测序。由于一位母亲在产后 6 个月回访时没有完成问卷调查,我们对其余 34 名婴儿根据 6 个月内是否出现湿疹进行分组,得到从未发生湿疹的 22 名健康婴儿(对照组)和 12 名湿疹婴儿(病例组)。
纳入、排除标准
出生方式、母体抗生素使用和母乳喂养是影响早期微生物群发展的首要影响因素,因此我们设计了严格的纳入和排除标准以尽量减少偏倚。本研究的纳入标准为:1)年龄在20-40岁之间的健康孕妇;2) 阴道分娩的婴儿;3) 孕妇孕前BMI < 28;4)孕妇具有纯母乳喂养的意愿;5)婴儿出生体重在2500-4000克之间。排除标准包括:1)妊娠期糖尿病、抑郁症、高血压等疾病;2) 分娩前3个月内服用过益生菌或抗生素;3) 分娩时使用抗生素;4) 孕期吸烟、饮酒史;5)分娩前3个月内腹泻;6)产后100天内使用抗生素;7) 在采样期间拒绝纯母乳喂养或主要母乳喂养;8) 诊断遗传性疾病。
最后,35 对母婴完成了4个时间点的纵向取样和同期的回访。一对母婴在第 6 个月时因未填写问卷而排除出组,最后获得34对母婴完整的配对样本及数据。在实验开始之前,所有参与者都已被告知实验存在的风险和参与者的权益,并且我们已获得每位受试者婴儿监护人签署的知情同意书。
伦理
所有与研究相关的方案均经南方医科大学珠江医院医学伦理委员会批准,并在 clinicalTrials.gov 注册,编号为NCT05462366。
临床研究人群和元数据
目前的队列包括 34 位阴道分娩的足月婴儿。没有使用统计方法来预先确定样本量,但是根据以往研究的样本量评估,我们假设 34 个婴儿的四个时间点样本足以说明微生物网络的动态变化情况。所有入组婴儿均在珠江医院妇产医学中心出生,出生后医生评估无任何疾病症状,并在最初24小时内进行母乳喂养。经过培训的医生负责收集胎便样本于无菌管中,并教导婴儿的父母在产后第 1、2 和 3 个月从洁净尿布中收集粪便样本到无菌管中的方法,并于每次采样前进行再次解说。收集样品后,将试管连同厌氧袋 (MGC AnaeroPack-Anaero) 一起保存在密封培养袋 (MGC AnaeroPack-Anaero) 中。培养袋和冰袋一同放入保温箱内,立即送至广东省科学院微生物国家重点实验室进行益生菌分离(如乳酸杆菌和双歧杆菌),其余样品液氮冷冻后-80℃保存直至制样。通过产后第 1、2、3 个月的回访和产后 第 6 个月的问卷调查收集母婴的健康状况、喂养方式、用药和一般信息。虽然婴儿大多是母乳喂养为主,但也有一些婴儿因某些原因偶尔会补充配方奶粉,包括母乳不足、进餐时间母亲不在婴儿身边、补充营养等。根据医生问诊的情况和问卷调查结果,我们将出生 6 个月内从未发生过湿疹的婴儿作为对照组,其余婴儿作为病例组。在所有参与者中,只有一名对照组的婴儿有湿疹家族史。
Roswall 等人的验证数据由来自原始队列随机选取的 16 名健康瑞典婴儿组成。在出生、第 4 个月、第 1 年和第 3 年收集的配对粪便样本16S rRNA扩增子测序数据用于网络分析。Christensen 等人的验证数据由来自原始队列随机选取的 16 名健康丹麦婴儿组成。在出生、1 个月和 1 岁时收集的配对粪便样本16S rRNA扩增子测序数据用于网络分析。
肠道菌群分析
按照制造商的说明进行样品制备,使用 E.Z.N.A.® Stool DNA 试剂盒(D4015,Omega, Inc., USA)从样品中提取 DNA。使用 341F (5'-CCTACGGGNGGCWGCAG-3') 和 805R (5'-GACTACHVGGGTATCTAATCC-3')引物,扩增了细菌和古细菌 16S rRNA 基因的 V3-V4 区域。16S rRNA 扩增子测序和生物信息学分析:根据 LC-Bio 的使用说明,使用 Illumina NovaSeq 平台对样品进行测序。使用 FLASH 合并双端测序序列。基于 QIIME2(2022-2版)使用 dada2 去噪(生成 ASV 表)、特征数据汇总、分类学分析(比对数据库为 Greengenes 13_8)和生成分类表。通过删除低丰度分类群(平均相对丰度 < 0.1%;亚组内出现率 < 10%)来过滤分类表。过滤后的分类表用于微生物分析。网络分析是基于 R 包 ggClusterNet(版本 0.1.0)进行的, MEN绘图基于model_igraph (method="cluster_fast_greedy") 函数模块。Alpha多样性、Beta多样性、主坐标分析(PCoA)、置换多元方差分析(PERMANOVA)是用自主开发的微生物分析 R 包 EasyMicroPlot(版本 0.5.1)进行的。
统计分析
所有数据分析均在 R 版本 4.2.1 中进行。为了探索网络拓扑学指数变化,我们使用 R 包 ggClusterNet(版本 0.1.0)基于 Spearman 相关性分析和 Benjamini-Hochberg 方法来调整 p 值。在网络分析中,所有分子网络都是基于 ASV 相对丰度的 Spearman 相关性构建的,Benjamini-Hochberg 调整的 p 值阈值为 0.05,相关系数的绝对值应大于 0.3。在统计分析之前,进行了Shapiro-Wilk 检验以验证测量的正态性。根据变量的分布,Mann-Whitney 检验或非配对的 Student's t检验用于计量资料统计。Kruskal-Wallis 平均值检验用于评估组间的非参数比较。使用单因素方差分析 (ANOVA) 检验多组之间差异的显著性,然后进行最小显著差数法 (Least Significant Difference, LSD) 进行多重比较。具有不同字母的组表示差异具有统计学意义,而具有相同字符的组表示在 LSD 检验中 p> 0.05。
贡献者
谢黎炜研究员和马颖教授设计了这项研究并获得了资助。马凯、王润鑫和王广贤代表江苏新申奥生物科技有限公司的成员,该公司提供横向经费支持该研究的开展。黄柳菁、盘桂花、冯译菲、范紫晶、黄广业、黄思霞、侯语汇和韩木兰参与样本的收集和测序数据的分析。黄柳菁和谢黎炜起草并编辑了文章,核对并验证了文章中报告的数据。
数据共享声明
所有 16S rRNA 基因扩增子测序数据均保存在国家微生物数据中心 (NMDC, https://nmdc.cn/) BioProjects NMDC10018172 下。来自 Roswall 等人和 Christensen 等人研究的验证 16S rRNA 基因扩增子测序数据分别列于补充表 4 和补充表 5 中。用于数据处理、分析和可视化的代码脚本已存放在 GitHub (https://github.com/xielab2017/EasyMicroPlot)。根据要求,可从相应的作者处获得更多信息。
致谢
我们非常感谢珠江医院妇产科的工作人员在志愿者入组及回访过程中提供的帮助与支持。感谢Xie Lab实验室的成员在样本采集和后续研究中提供的出色协助。以及为本研究提供资助的江苏新申奥生物科技有限公司和中国国家自然基金以及广东省基础与应用基础杰出青年基金的支持。
引文格式:
Huang, Liujing, Guihua Pan, Yifei Feng, Zijing Fan, Kai Ma, Runxin Wang, Guangxian Wang, Guangye Huang, Sixia Huang, Yuhui Hou, Mulan Han, Liwei Xie, and Ying Ma 2023. “Microbial network signatures of early colonizers in infants with eczema.” iMeta. e90. https://doi.org/10.1002/imt2.90
-
点赞 (0人)
- 收藏 (0人)

