
iMeta | 农科院资划所刘卫星组揭示氮富集条件下土壤细菌性状介导网络复杂性的机制
- 看不见的线
- 5044
- 2024-06-07 15:31:50
- 文章来源:iMeta
长期氮输入通过改变生活史策略降低温带草原土壤细菌网络复杂性

iMeta主页:http://www.imeta.science
研究论文
●原文链接DOI: https://doi.org/10.1002/imt2.194
●2024年4月15日,中国农业科学院农业资源与农业区划研究所刘卫星团队在iMeta在线发表了题为 “Long-term nitrogen input reduces soil bacterial network complexity by shifts in life history strategy in temperate grassland” 的研究文章。
●本文通过研究不同氮输入梯度下土壤细菌和真菌群落的网络结构以及微生物性状,发现土壤细菌生活史策略的改变调控其群落共现网络复杂性的降低,揭示了微生物种间相互作用和共现网络的性状调节机制。
●第一作者:王超
●通讯作者:刘卫星(liuweixing@caas.cn)
●合作作者:石紫阅、李澳归、耿天懿、刘玲莉
●主要单位:中国农业科学院农业资源与农业区划研究所、中国科学院植物研究所
亮 点
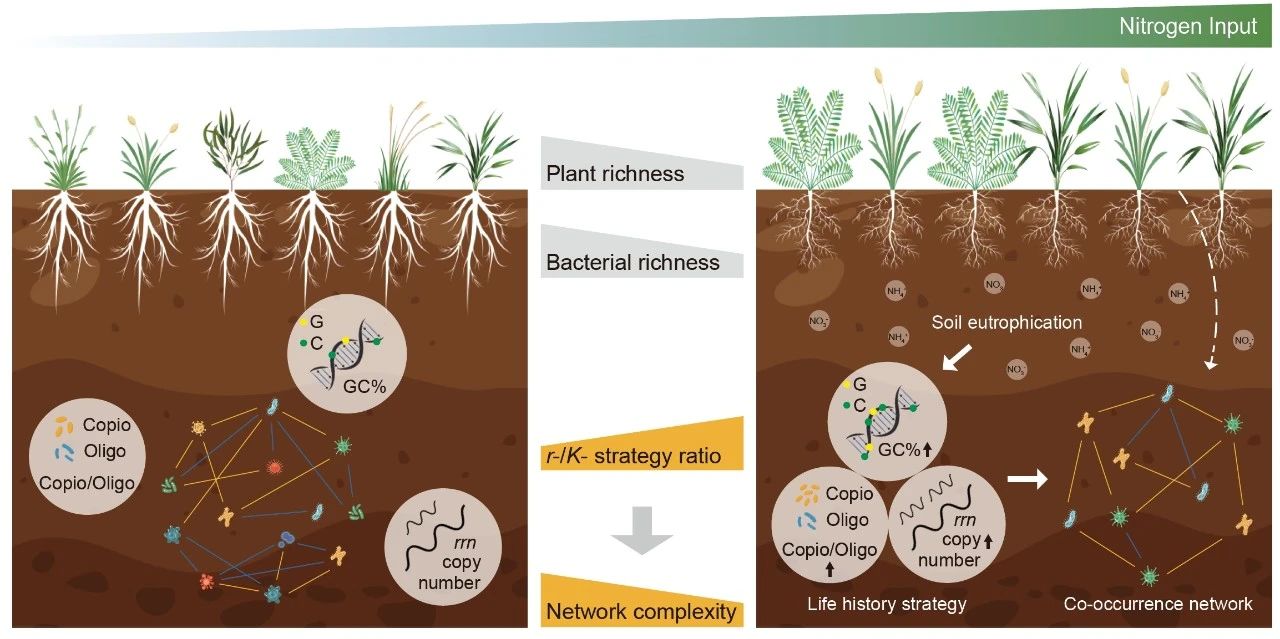
● 土壤细菌共现网络复杂性和负/正凝聚力比值均随施氮量的增加而降低;
● 氮素富集导致土壤细菌群落趋向富营养型, r-/K-策略比例增加;
● 氮富集条件下r-/K-策略比的增加驱动细菌共现网络复杂性的降低。
摘 要
通过研究不同氮输入梯度下土壤细菌和真菌群落,构建共现网络,并评估微生物性状。结果表明,土壤细菌共现网络的复杂性随着施氮量的增加而降低。随着施氮量的增加,负凝聚力与正凝聚力之比下降,表明竞争性减弱,但合作性相互作用增强。然而,土壤真菌网络的复杂性在氮富集下并没有改变。此外,氮输入促进土壤细菌的富营养型/寡营养型比率、核糖体RNA操纵子(rrn)拷贝数和鸟嘌呤-胞嘧啶(GC)含量,随着r-/K-策略比的增加,细菌生活史策略向富营养型转变。分段结构方程建模结果进一步表明,细菌共现网络复杂性的降低是由细菌r-/K-策略比的增加直接调控的,而不是细菌丰富度的降低。我们的研究揭示了微生物性状在全球变化下调节相互作用和塑造共现网络的机制。
视频解读
Bilibili:https://www.bilibili.com/video/BV1ft421w79V/
Youtube:https://youtu.be/gI94G-Yrd8w
中文翻译、PPT、中/英文视频解读等扩展资料下载
请访问期刊官网:http://www.imeta.science/
全文解读
引 言
化石燃料燃烧和氮肥施用排放的含氮(N)化合物导致N沉降量已经增加200%,预计到2050年全球各地区将增加一倍。N沉降造成的N富集对植物多样性、群落和陆地生态系统(特别是草原)的功能构成严重威胁。生态系统功能不仅与植物的变化有关,还与土壤微生物响应N输入的动态有关。微生物种群的改变可以引发微生物之间相互作用的变化,这在维持微生物网络的复杂性、稳定性和功能方面发挥着重要作用。因此,了解微生物的共现和相互作用对于预测草原生态系统中N富集的生态系统过程和功能至关重要。
微生物相互作用由合作和竞争组成,通常与丰富度相关。然而,最近一些关于N富集对微生物相互作用的研究具有不同的结果,这与微生物丰富度如何变化无关。具体来说,一些研究报告施N后土壤细菌相互作用和网络复杂性增加,这归因于关键类群数量的增加。相反,对150年N肥施用的调查显示,土壤细菌网络的复杂性显著降低,同时群落内合作关系也有所下降。此外,一项多梯度N添加研究表明,在较低的N输入条件下,细菌网络复杂性较高,但在较高的N输入条件下,复杂性降低,并且合作关系趋势相同。在这些研究中,发现合作关系对N输入的响应与细菌网络复杂性的变化一致。
与合作对微生物网络复杂性的调节作用不同,一些群落生态学家提出,竞争性相互作用的存在能促进高阶相互作用,从而增强共现网络的复杂性和稳定性。即竞争性相互作用的增强有助于增加微生物网络的复杂性。相应地,上述关于合作减少并伴随网络复杂性简化的结果,似乎与正(负)相互作用和复杂性之间的理论共存概念陈述相矛盾。因此,尽管对全球草地生态系统中的微生物多样性和群落进行了广泛的调查研究,但关于微生物相互作用和网络复杂性如何响应N富集及其潜在机制的信息仍然严重缺乏。
在这里,我们研究了中国温带草原微生物共现网络随N输入梯度的变化。本研究旨在探讨土壤微生物相互作用对N输入的响应,并阐明潜在的驱动因素和机制。我们将解决以下问题:(1)在N富集条件下,微生物丰富度的降低是否会导致微生物网络复杂性的降低?(2)微生物网络复杂性中的合作(竞争)相互作用的变化如何响应N富集?(3)考虑到土壤真菌与植物的联系更为密切,N输入对真菌网络复杂性的影响是否更明显?
结果与讨论
试验地点位于中国内蒙古多伦县半干旱草原。自2003年起采用拉丁方设计,总共包括8个N添加水平(0、1、2、4、8、16、32和64 g N m−2 y−1)。土壤样本于2016年8月采集并用于测序和化学分析。随着N输入的增加,细菌网络特征的节点数和边数、平均度、聚类系数和网络拓扑参数主成分分析第一轴(PC1)均呈现显著的递减趋势(图1A、B和S1)。与拓扑参数类似,细菌群落的总凝聚力、正凝聚力、负凝聚力以及负/正凝聚力随着N输入的增加而减少(图1C)。然而,N输入并没有改变土壤真菌网络的复杂性(图S2)。随着N输入的增加,富营养/寡营养细菌比率、核糖体RNA操纵子(rrn)拷贝数、鸟嘌呤-胞嘧啶(GC)含量和代表细菌r-/K-策略比率的PC1值逐渐增加(图2A)。总凝聚力与细菌丰富度呈正相关,但与r-/K-策略比率呈负相关(图2B)。分段结构方程模型(SEM)结果表明,N输入通过增加土壤溶解性无机氮含量(DIN)和降低土壤pH值,改变细菌r-/K-策略比率。细菌r-/K-策略比率对细菌丰富度产生负面影响。细菌总凝聚力与r-/K-策略比率呈直接负相关,但与细菌丰富度没有相关性(图2C)。
与高N条件下更大的rrn拷贝数一致,我们发现N输入促进土壤DIN并增加细菌rrn拷贝数向富营养群落发展(图S3)。我们进一步发现GC含量随着施N量增加而增加。根据分子生物合成规则,每个GC碱基对需要8个N原子,而腺嘌呤-胸腺嘧啶(AT)碱基对只需要7个N原子。“资源驱动选择” 理论提出,N限制是一种强有力的选择作用,将导致GC含量相对较低。因此,较高的土壤N含量将通过缓解N限制而导致核苷酸序列具有G+C偏倚。与以往报道不同的是,区域尺度上GC含量与土壤pH呈正相关,我们的研究中GC含量与土壤pH呈负相关(图S4)。这可能是由于额外施N导致土壤pH值下降所致。总体而言,较高的细菌r-/K-策略比率表明细菌生活史策略倾向于r-选择策略。受到促进的r/K策略比率支持温带草原细菌群落响应N富集的 “资源驱动选择”。
高N输入下细菌网络复杂性的降低伴随着丰富度的降低,这在我们之前的工作中发现,并呈现出正相关关系(图2B)。我们发现丰富度的降低归因于寡营养类群的丧失(图S5),这导致生活史策略的转变。当控制r-/K-策略比率时,总凝聚力与丰富度的偏回归相关性不再显著(图2B)。此外,SEM结果还表明,细菌群落复杂性与r-/K-策略比率呈负相关,这意味着细菌连接的紧密程度是由生活史策略驱动的,而不是细菌丰富度(图2B、C、和S6)。寡营养类群通常从各种复杂的顽固物质中获取营养或能量。这一获取过程涉及多种微生物参与的多种代谢途径,在微生物类群之间建立更有效、更强的复杂联系。然而,N输入促进植物生长和随之而来的新鲜碳输入(图S7),产生足够的能量供应和活性的底物。过量的N以及底物的应用有利于富营养型微生物并促进细菌的独立性,削弱复杂的连接并导致连通性减少,从而简化共现网络。r-/K-策略比率介导细菌共现网络复杂性这一发现,指出细菌遗传和编码性状在分类群共现中的调节作用。
我们推测凝聚力的降低与群落中高阶相互作用的解构有关。相互作用链是指一个物种通过除成对相互作用之外第三个物种的丰度变化而对另一个物种产生的间接影响。这些出现的相互作用链可以产生高阶相互作用,获得更多样化的成对关系,并维持复杂而稳健的共现网络。凝聚力的降低和细菌共现网络的简化反过来在某种程度上表明,在N富集条件下,高阶相互作用的解构以及向低阶相互作用的转变。
负凝聚力与正凝聚力比率代表竞争或合作强度的变化。比率越高表明群落内类群之间的竞争越激烈。与典型草原中负/正凝聚力比率增加相反,我们的结果表明,随着N输入的增加,负/正凝聚力比率降低。这表明群落内竞争的减少和合作的加强。具体来说,额外的N输入能直接减轻微生物之间对养分的竞争,并可能削弱竞争相互作用。此外,不同类群内部的快速生长和代谢会产生代谢物,促进类群之间的能量和物质交换,称为交叉饲喂,以增强合作。相应地,我们发现随着细菌r-/K-策略比率的增加,合作增强(图S8)。此外,某些竞争可以通过均衡群落内其他类群的适应性来促进共存。此类竞争对手的流失可能会导致其他竞争对手的后续流失以及共现网络的简化。同时减少的竞争相互作用和简化的网络强调竞争在维持物种共存中的作用。
土壤真菌共现网络复杂性不变的发现并不支持我们的第三个假设,即土壤真菌对N输入的响应更灵敏。未改变的共现网络可能归因于土壤真菌的生理和形态特性。土壤真菌通常具有很强的矿化复杂有机化合物以获取N的能力。此外,它们在生物量积累中也具有高N利用效率和高C/N比。这两个特性有利于土壤真菌在低N条件下生长,对额外N输入的依赖性较小,表现出对N输入不敏感。此外,真菌较厚的细胞壁和内在孢子对土壤酸化和金属毒性表现出高度耐受性,能在N富集下保持稳定。真菌菌丝体的特性使其能够穿透并扩散到地下,建立菌丝网络。因此,土壤真菌菌丝网络可以延伸分布到更广泛的空间,从而能够抵抗环境变化。
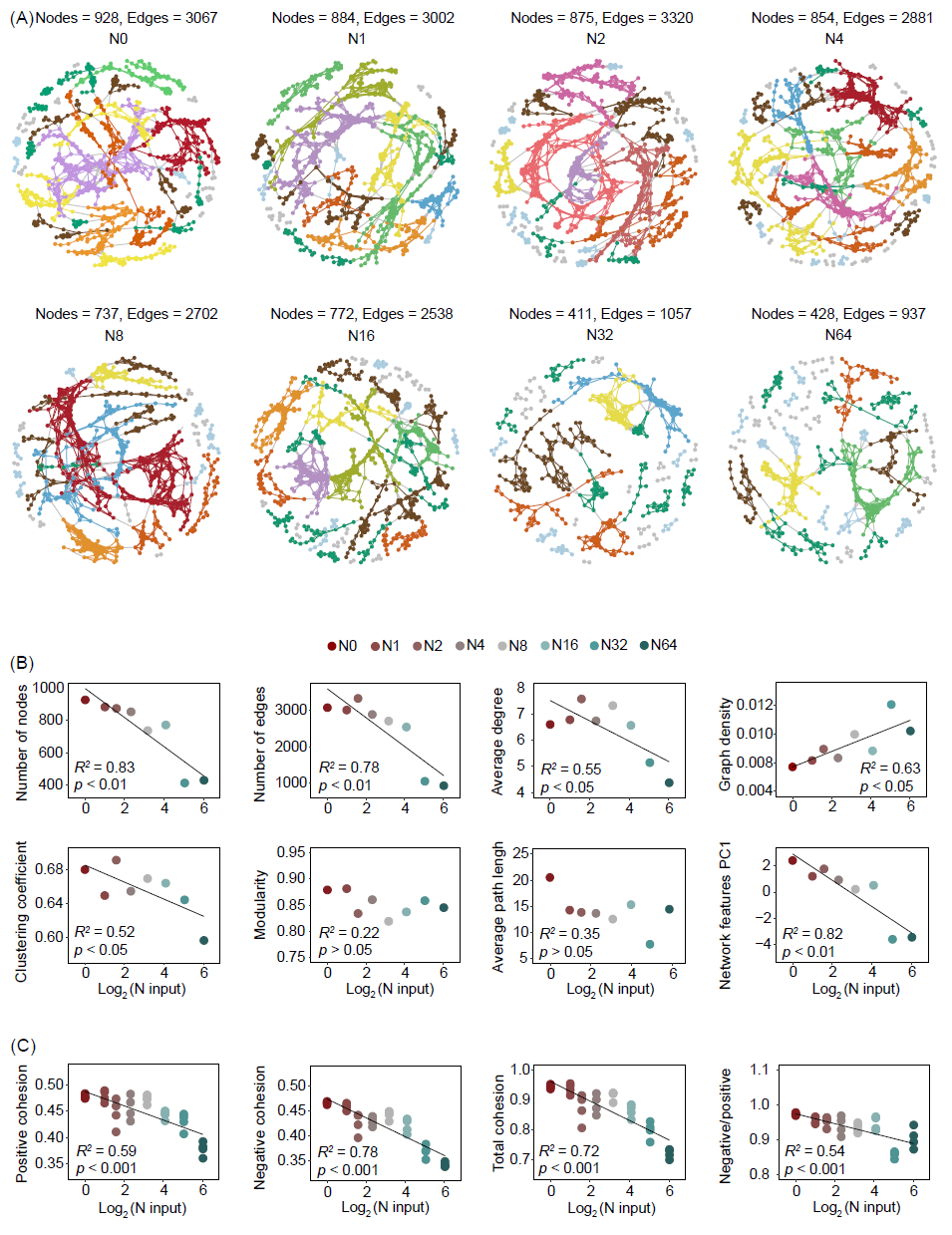
图 1. 不同氮(N)输入处理下的细菌共现网络、网络拓扑参数和凝聚力
(A)不同N输入处理下的细菌共现网络;(B)拓扑参数包括节点数、边数、平均度、图密度、聚类系数、模块度、平均路径长度和上述拓扑参数主成分分析第一轴的值(PC1);(C)不同N输入下的土壤细菌共现网络的正凝聚力、负凝聚力绝对值、总凝聚力和负/正凝聚力绝对值。节点表示各个操作分类单元(OTU),而边表示OTU之间的显著相关性。节点的颜色根据模块中节点的数量进行分类。网络特性是拓扑参数的主成分分析的PC1值。拓扑参数包括节点数、边数、平均度、图密度、聚类系数、模块度和平均路径长度。图中仅展示显著的线性回归。
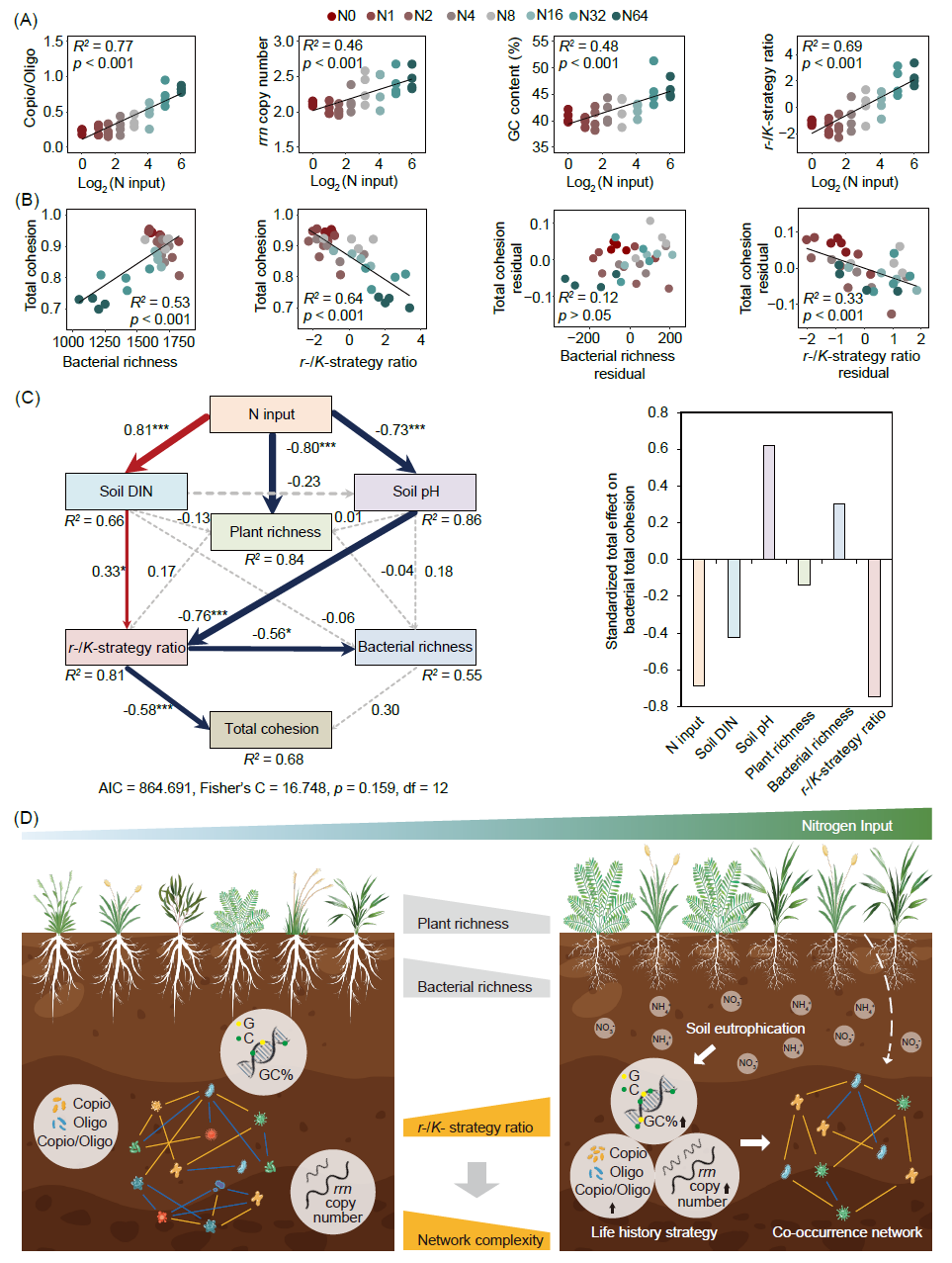
图 2. 氮(N)输入处理下细菌的生活史特性及细菌网络复杂性的影响因素
(A)随着N输入的增加,土壤细菌富营养/寡营养比率、核糖体RNA操纵子(rrn)拷贝数、鸟嘌呤胞嘧啶(GC)含量以及细菌r-/K-策略比率;(B)总凝聚力和丰富度与r-/K-策略比之间的关系,控制r-/K-策略比时,细菌丰富度残差和总内聚力残差的偏回归;(C)分段结构方程模型(SEM)用于检验细菌总凝聚力对N输入的响应的预测因素和途径。细菌r-/K-策略比率是主成分分析第一轴的值(PC1),包括富营养/寡营养比率、rrn拷贝数和GC含量。SEM的变量包括土壤溶解无机氮(DIN)、土壤pH、植物丰富度、土壤细菌r-/K-策略比率和细菌丰富度。红色和蓝色实心箭头表示统计显著(p < 0.05)正相关和负相关。红色和蓝色箭头的宽度与标准化路径系数的强度成正比。*和***分别表示p < 0.05和p < 0.001。R2表示解释方差的比例。模型的评估参数是Fisher's C(p > 0.05)。(D)概念图描绘了氮输入如何通过调节细菌生活史策略而不是通过减少植物或细菌丰富度来影响细菌共现网络的复杂性。土壤富营养化导致细菌富营养/寡营养比率、rrn拷贝数和GC含量增加,从而使细菌群落转向富营养策略。细菌生活史策略的这种转变有助于降低细菌共现网络的复杂性。虚线表示植物丰富度对细菌网络的影响不显著。“Copio/Oligo”指富养型/寡养型比率,“GC%”指GC含量。图中仅展示显著的线性回归。
结 论
总之,我们的研究表明,在温带草原中,随着N输入的增加,细菌和真菌网络以及相互作用具有明显不同的模式。随着N输入的增加,细菌网络复杂性呈现下降趋势,伴随着负/正凝聚力比率下降,而真菌群落的网络复杂性保持不变。我们进一步发现,细菌网络复杂性的下降是由N富集下细菌r-/K-策略比率的增加驱动的。值得注意的是,这个结果是基于十多年的N输入研究得出的。考虑到施氮量随时间的累积效应,在外推时应考虑土壤细菌网络复杂性响应N输入的时间动态。尽管如此,据我们所知,这项研究是第一份实证报告,表明细菌共现网络复杂性的降低主要归因于草原生态系统N富集下细菌生活史策略特性的变化。这一发现能为更全面地理解微生物性状调节共现网络和相互作用以响应环境变化的潜在机制提供见解。
方 法
实验设计
试验地点位于中国内蒙古多伦县半干旱草原(42.01′ N,116.16′ E,海拔1324 m)。该地区受季风影响,冬季寒冷干燥,夏季温暖潮湿。年平均气温2.1 ℃,月平均气温-17.5 ℃(1月)至18.9 ℃(7月)。年平均降水量382.3 mm,7、8月约占50%。根据联合国粮食及农业组织的分类,土壤类型为Haplic Calcisols,含砂粒69.21±0.06%,粉粒15.60±0.02%,粘粒15.19±0.02%。土壤有机碳和总氮(N)浓度为16.94 ± 2.34 g kg−1分别为1.65±0.27 g kg−1,土壤pH值为6.84 ± 0.02,环境N沉降量约为1.6 g N m−2。植物群落以多年生草本植物和禾本科植物为主,包括Stipa krylovii Roshev.,Agropyron cristatum L.,Artemisia frigida Willd和Cleistogenes squarrosa Trin.。
自2003年开始进行N肥试验,采用拉丁方设计,试验小区64块,面积10 × 15 m。这些地块排列成8行8列,相邻地块之间有5 m的缓冲区。每行有8个地块,随机分配给8个N施用水平之一(0、1、2、4、8、16、32和64 g N m−2 y−1 )。每年七月初,N以尿素的形式施用。2005年起,8月下旬对地面进行四行割草(每两行割一行),并清除试验地块内的干草。在这项研究中,样本完全从未进行割草的地块中收集。
土壤化学性质和植物调查
土壤样品采集于2016年8月15日,用土钻在每个小区内随机采集6钻土芯(深15 cm,直径5 cm),混合为一份样品。将根和石头通过2 mm的筛子筛分后,土壤样品储存在冰盒中并立即转移到实验室。将土壤样品分为两份,其中一份储存在4 ℃冰箱用于土壤理化分析,另一份保存在-80 ℃冰箱用于DNA提取。使用玻璃电极对土水比为1:2.5的悬浮液测定土壤pH值。使用2 M KCl溶液提取土壤溶解的无机N(DIN),并用流动分析仪测定。
采用点截距法对各样地1 × 1 m永久样方内生物量峰值时的植物群落进行调查。通过在永久样方上放置1 × 1 m框架和100个均匀间隔的10 × 10 cm网格来估计植物物种丰富度。然后将每个实验地的植物物种丰富度记录为样方中存在的植物物种数量。
扩增子测序
使用PowerSoil DNA分离试剂盒(MoBio Laboratories)提取土壤DNA,每个样品称重0.5 g新鲜土壤,并遵循制造商的说明进行操作。细菌群落使用引物338F(5'-ACTCCTACGGGAGGCAGCAG-3')和806R(5'-GGACTACHVGGGTWTCTAAT-3')来扩增16S rRNA基因的V3-V4高变区。对真菌群落内部转录间隔区(ITS1)使用ITS1-F(5'-CTTGGTCATTTAGAGGAAGTAA-3')和ITS2(5'-TGCGTTTCTTCATCGATGC-3')引物进行扩增。将每个样品独特的10 bp条形码附在引物的5'端,以区分每个样品。50 μL PCR反应体系包含 5 μL 10× Ex Taq 缓冲液(Mg2+ plus)、4 μL 12.5 mM dNTP Mix、1.25 U Ex Taq DNA聚合酶(TaKaRa)、2 μL模板DNA和36.75 μL ddH2O。PCR混合物在94 °C预变性2 min,然后进行30个扩增循环(94 °C 30 s,57 °C 30 s,72 °C 30 s),最后在72°C延伸10 min。每个样品进行3次PCR,每批均设有阴性对照。PCR产物以等摩尔浓度混合并使用QIAquick凝胶提取试剂盒(Qiagen)纯化。纯化的PCR产物在Illumina MiSeq平台(Illumina)上进行测序。
原始读数被修剪,低质量碱基(Phred score < 20)被删除。使用FLASH软件(version 1.0.0)根据每个样本的唯一条形码进行序列拆分。使用MOTHUR软件从这些序列中去除条形码和引物。将剩余序列聚类成具有97%相似度的操作分类单元(OTU),并在此过程中去除嵌合体。采用Ribosomal Database Project(RDP)分类器,基于SILVA 128数据库和UNITE fungal ITS 7.2数据库对16S rRNA和ITS基因序列进行分类类群划分,鉴定阈值为70%。同时,未分类的细菌和真菌OTUs也被去除。并将所有细菌(18,877)和真菌(24,945)样品分别稀疏抽平到相同的深度。所有序列已存入NCBI’s SRA数据库,项目号为PRJNA573484和PRJNA573488。
数据分析
所有统计分析均使用R 4.3.1,数据可视化使用“ggplot2”包。在不同的N输入处理下,分别构建细菌和真菌群落共现网络。在相同的N输入处理中,只有相对丰度大于0.01%且存在于所有样本中的OTUs才包含在网络中。计算Pearson相关系数以确定OTUs之间的相关性,这些相关性用于表示网络中的边。为了最大限度地减少网络构建和比较中的不确定性,使用“RMThreshold”包根据随机矩阵理论确定微生物网络构建阈值,识别阈值为0.75,并选择p值小于0.01的边。
微生物相互作用和复杂性是通过构建复杂的共现网络来评估的,共现网络包含正相关和负相关,代表共存类群之间的合作和竞争行为。网络中节点的数量代表网络中存在的物种的数量。使用“igraph”包可视化细菌和真菌群落共现网络,并计算网络拓扑参数,包括节点数、边数、平均度、聚类系数、图密度、平均路径长度和模块化。这些参数用于评估共现网络的复杂性。采用双变量线性回归分析细菌群落和真菌群落的拓扑参数与N输入水平之间的关系。N输入量经过对数转换。对拓扑参数进行主成分分析,然后使用主成分分析第一轴(PC1)的值来表示细菌和真菌网络特性和复杂性。提取正相关边和负相关边,并进行N输入水平下的回归分析。
在之前的方法的基础上,计算凝聚力来表示细菌群落的复杂性。公式如下:
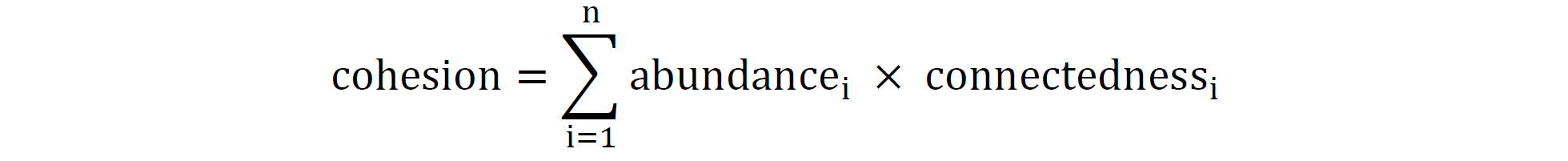
其中n代表群落中类群的总数。通过对类群之间显著的正相关性和负相关性进行求和,并按各自的丰度进行加权,来测量每个样本的正凝聚力值和负凝聚力值。负凝聚力和正凝聚力的值范围分别为-1到0和0到1。总凝聚力绝对值越高表示相关性越高,负凝聚力绝对值越大表示稳定性越高。总凝聚力通过将正值和负凝聚力绝对值相加来确定。使用绝对值测量负凝聚力和正凝聚力之间的比率来表示竞争或合作强度的变化。还使用二元线性回归来分析凝聚力指数与N输入水平之间的关系。
与生活史相关的细菌特性根据三个参数进行量化。根据现有的生活史策略分类,鉴定富营养型(r策略)和寡营养型(K策略)细菌。其中,α-Proteobacteria、δ-Proteobacteria、Plantomycetes和Acidobacteria可以矿化顽固性碳作为碳(C)资源和能量生长,Chloroflexi生长缓慢,在养分不足的土壤中生长旺盛。所有这些类群都被归类为具有K策略的寡营养细菌。相反,由于使用不稳定的C底物,β-和γ-Proteobacteria以及Bacteroidetes被定义为富营养细菌。此外,由于对新鲜有机物的分解,Firmicutes也被认为是富营养细菌。计算这两个策略(富营养/寡营养)的相对丰度比例来代表细菌生活史策略的变化。
根据rrn DB数据库(version 5.8,https://rrndb.umms.med.umich.edu/)估计细菌核糖体RNA操纵子(rrn)在群落水平的平均拷贝数。我们从最低等级(species)开始匹配rrn拷贝数。如果最低级别没有匹配,则使用较高级别的平均rrn拷贝数。然后通过所有OTU的丰度加权平均rrn拷贝数计算群落水平的rrn拷贝数。
通过将16S rRNA代表序列注释到基因组分类数据库(Genome Taxonomy Database, GTDB)中,计算群落水平细菌平均GC含量。我们下载了在一组细菌代表性基因组中鉴定的16S rRNA基因序列的FASTA文件(https://data.gtdb.ecogenomic.org/releases/latest/genomic_files_reps/bac120_ssu_reps.tar.gz),并通过makeblastdb工具将其转换为参考数据库。使用BLAST(version 2.15.0)将16S代表性序列与参考数据库比对,一致性 ≥ 97%。获取登录ID后,GC含量与GTDB中下载的元数据文件进行匹配(https://data.gtdb.ecogenomic.org/releases/latest/bac120_metadata.tsv.gz)。然后通过所有OTU的丰度加权平均GC含量计算群落水平的GC含量。然后根据上述三个性状进行主成分分析,用PC1值代表细菌r-/K-策略比。
采用分段结构方程模型(Piecewise structural equation modeling, SEM),利用“piecewiseSEM”包来研究N输入调节细菌性状和网络复杂性的直接和间接途径。该模型提出了土壤特性、植物丰富度、细菌丰富度和性状之间假设的因果关系。基于富营养/寡营养比率、rrn拷贝数和GC含量,将细菌性状量化为r-/K-策略比率。Fisher's C统计量用于评估SEM的拟合优度。依次从初始SEM中删除不重要的路径,并根据最低的Akaike Information Criterion(AIC)选择最终SEM。
数据可用性声明
16S和ITS序列已提交到NCBI数据库,登录号为PRJNA573484(https://www.ncbi.nlm.nih.gov/search/all/?term=PRJNA573484)和PRJNA573488(https://www.ncbi.nlm .nih.gov/search/all/?term=PRJNA573488)。使用的数据、脚本和表格保存在GitHub(https://github.com/ChaoWang-01/code-for-imeta)中。补充材料(方法、图、脚本、图文摘要、幻灯片、视频、中文翻译版本和更新材料)可以在在线DOI或iMeta Science中找到http://www.imeta.science/。
引文格式:
Wang Chao, Ziyue Shi, Aogui Li, Tianyi Geng, Lingli Liu, Weixing Liu. 2024. Long-term nitrogen input reduces soil bacterial network complexity by shifts in life-history strategy in temperate grassland. iMeta 3: e194. https://doi.org/10.1002/imt2.194
-
点赞 (0人)
- 收藏 (0人)

