
【文献速递】中国鲍曼不动杆菌血流分离株的基因组流行病学与系统动力学
- 看不见的线
- 2796
- 2025-08-19 08:22:03
- 原创

文章题目:Genomic epidemiology and phylodynamics of Acinetobacter baumannii bloodstream isolates in China
文章链接:https://doi.org/10.1038/s41467-025-58772-9
发表时间:2025.4
发表期刊:Nature communications
IF:15.7
研究背景
鲍曼不动杆菌是一种机会性病原体,主要引起医院获得性感染,如呼吸机相关性肺炎和血流感染(BSIs),尤其在重症患者中常见。全球每年约有100万人感染鲍曼不动杆菌,其血流感染的死亡率超过60%,多重耐药(MDR)比例约为45%。在中国,CRAB的分离率高达约70%,仅对多黏菌素和替加环素敏感。同时鲍曼不动杆菌还具有很强的环境适应性,包括消毒剂、干燥耐受性,生物膜形成能力和运动性,能在医院环境中长期存活。此外,传统认为鲍曼不动杆菌毒力较低,但近些年研究却显示其毒力逐渐增强。
本研究通过对中国2011-2021年间1506株鲍曼不动杆菌血流感染分离株的基因组分析,揭示了中国鲍曼不动杆菌的种群结构、时空分布和进化动态。重点探讨了ST208谱系的崛起原因,包括其毒力增强、耐药性积累、环境适应性和基因组可塑性。
研究思路
1、通过全国范围的菌株收集和全基因组测序建立基础数据库,运用Oxford MLST分型和Kaptive荚膜分型进行分子特征鉴定。
2、结合表型实验(如补体杀伤实验、小鼠感染模型和干燥耐受实验)与耐药基因、毒力基因分析,系统评估不同菌株的生物学特性。
3、基于比较基因组学分析(包括重组、SNP和移动遗传元件)分析生物学特性差异的可能原因。
4、采用贝叶斯系统地理学方法重建菌株传播历史,并通过离散性状分析量化跨区域传播事件。
研究结果
1、种群结构及时空分布
2011-2021年间,作者从25个省份的76家哨点医院共收集了1506株参与血流感染(BSI)的非重复鲍曼不动杆菌分离株。菌株最多的三个省份是浙江省(ZJ)(40.2%)、安徽省(AH)(20.0%)和山东省(SD)(7.7%)。
①物种鉴定:通过平均核苷酸同一性(ANI)和核糖体多位点序列分型(rMLST)对基因组进行分析,排除了钙醋酸不动杆菌-鲍曼不动杆菌(Acb)复合体的其他细菌物种。
② ST型:通过MLST (Oxford)分型方案,共鉴定出149种ST型,其中有47种是作者新提交的ST型((ST3225-ST3271)。数量排名前三的ST是ST195(n = 334)、ST208(n = 234)和ST191(n = 148)。81.74%的菌株(1231/1506)属于IC2,其核心节点为ST208。
③荚膜分型:共鉴定出101种KL分型,排名前3位的是KL3(22.2%)、KL2(11.6%)和KL9(9.6%),表明中国引起BSI的鲍曼不动杆菌分离株中KL型高度多样化。
④脂多糖分型:共鉴定出16种OCL分型,排名前3位的是OCL1(83.1%)、OCL6(7.2%)和OCL2(3.9%)。
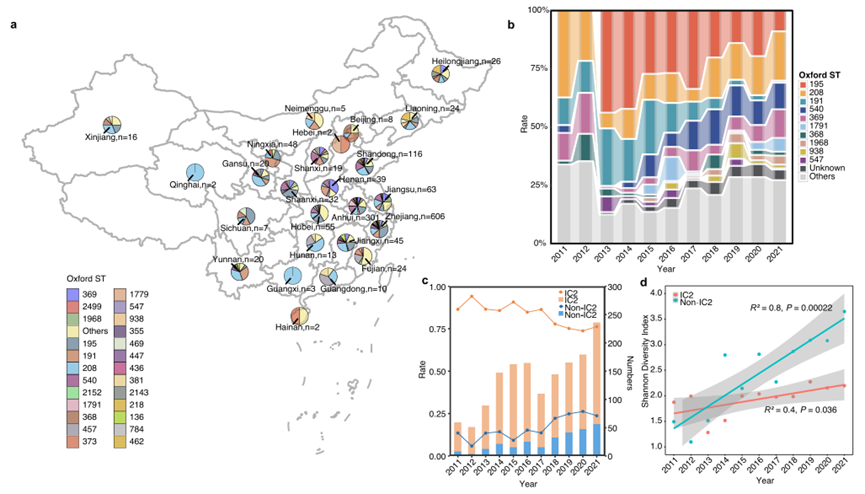
图1.中国BSI鲍曼不动杆菌分离株的种群结构、动态和时空分布。(a.分离株的地理位置;b.分离株ST的历年分布情况;c. IC2和非IC2菌株的数量和比例变化;d. IC2和非IC2菌株的Shannon多样性指数)
2、种群动态及系统发育关系
① IC2的ST变化趋势:ST195和ST191的大幅下降和ST208的显著上升。(ST195从42.18%降至8.5%,ST191从18.37%降至0.9%,ST208从12.93%升至21.19%)
②中国与全球的ST分布对比:截至2023年9月,于NCBI Pathogen Detection数据库筛选到1978株全球BSI分离株(含中国大陆246株),时间跨度1995–2023年。中国与全球仅共享30种ST(共207种),前三大STs(ST195、ST208、ST191)相同,但其余Top 10 ST完全不同,表明中国大陆与全球其他地区的BSI鲍曼不动杆菌ST分布存在显著差异。
③ ST208的进化优势:cgMLST显示1506个分离株可分为IC2(紧密聚类)和非IC2(分散分布)两个簇。SNP最大似然树显示,IC2进一步分为四大谱系:ST191、ST208、ST195和其他STs。然后ST208又可以细分为3个主分支(L1-L3),L3又可以分为11个亚分支。ST208的不同KL(KL2和KL7)在树上交叉分布,表明不同的 KL可能是通过重组获得的。L1主要是从2016年或之前分离出来(36/44, 81.82%),全部L2(n = 22)和大部分L3(102/159, 64.15%)是在2017年及之后分离出来的。结果表明ST208-KL7已经进化成一个独特的簇,可能进一步扩大并逐渐取代ST208-KL2或与它共存。
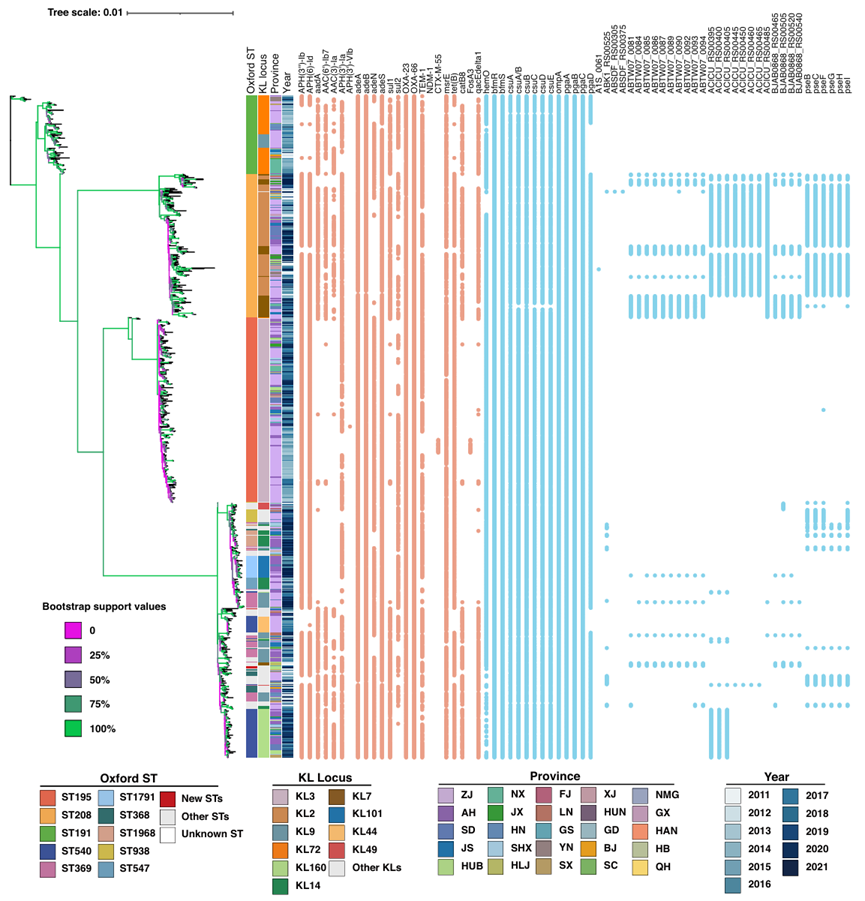
图2. IC2 BSI鲍曼不动杆菌分离株的最大似然系统发育,以及代表性的抗菌素耐药基因和毒力基因。
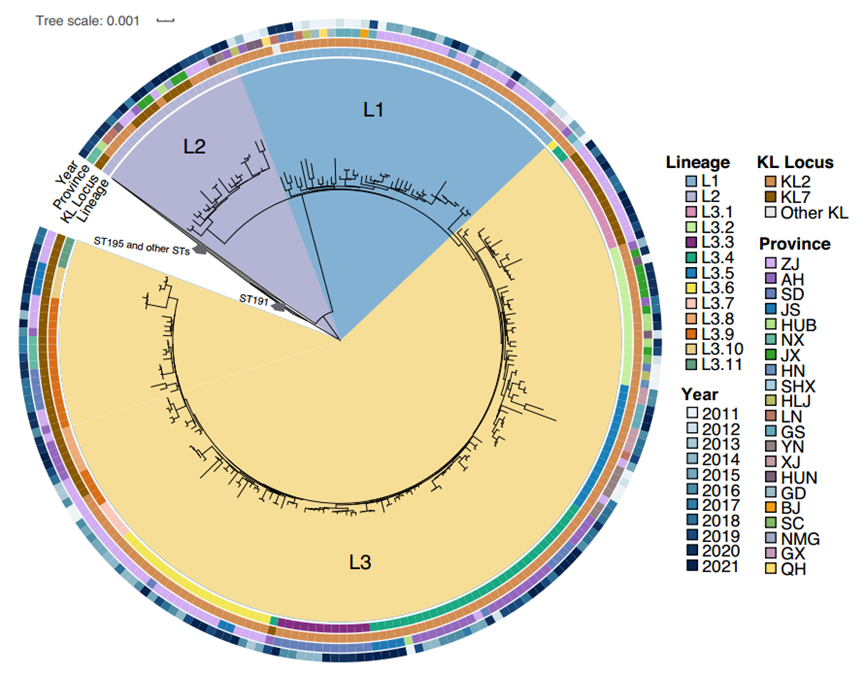
图3. ST208谱系在IC2最大似然系统发育树中的细分。
3、毒力研究
比较ST208与ST191、ST195的毒力差异,重点关注荚膜(capsule)的作用。根据地区和分离时间,每组ST-KL组合随机选取5株菌进行实验。
①补体杀伤实验(体外):ST208(尤其是ST208-KL2)对血清补体的抵抗力最强。
②小鼠感染模型(体内):ST208菌株毒力最强,感染ST208-KL2或ST208-KL7的小鼠存活率显著低于ST191-KL72或ST195-KL3,但是ST208、ST191、ST195的平均毒力仍弱于经典高毒株LAC-4和AB5075。
③毒力基因数量差异:ST208携带的毒力基因数量最多(133.90±4.72),显著多于ST191(126.44±6.86)和ST195(131.79±1.61)。
④毒力基因PCA分析:ST191、ST195和ST208的毒力基因存在显著差异,其中ST195的毒力基因谱与其它两者的差异最大。
⑤毒力基因的时空动态:免疫调节相关基因(如T6SS、hemO簇)在ST208中富集,在IC2中的携带率显著高于非IC2,可能是其成为主导克隆的关键。
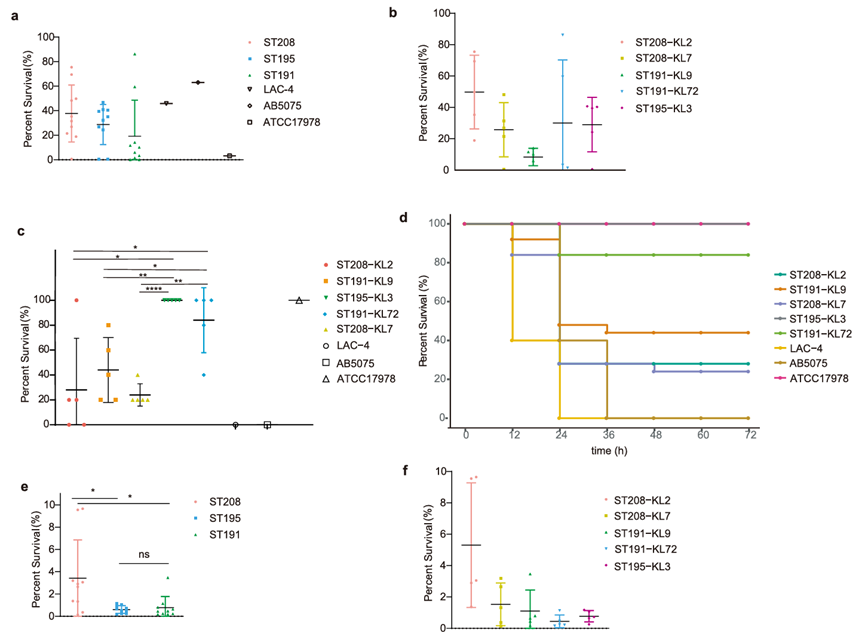
图4 ST208、ST191和ST195分离株的毒性和干燥耐受性的实验研究结果。(a-b.不同ST或ST-KL菌株的补体杀伤试验;c-d.不同ST或ST-KL菌株的小鼠感染模型存活率;e-f.不同ST或ST-KL菌株在20%湿度下的存活率)
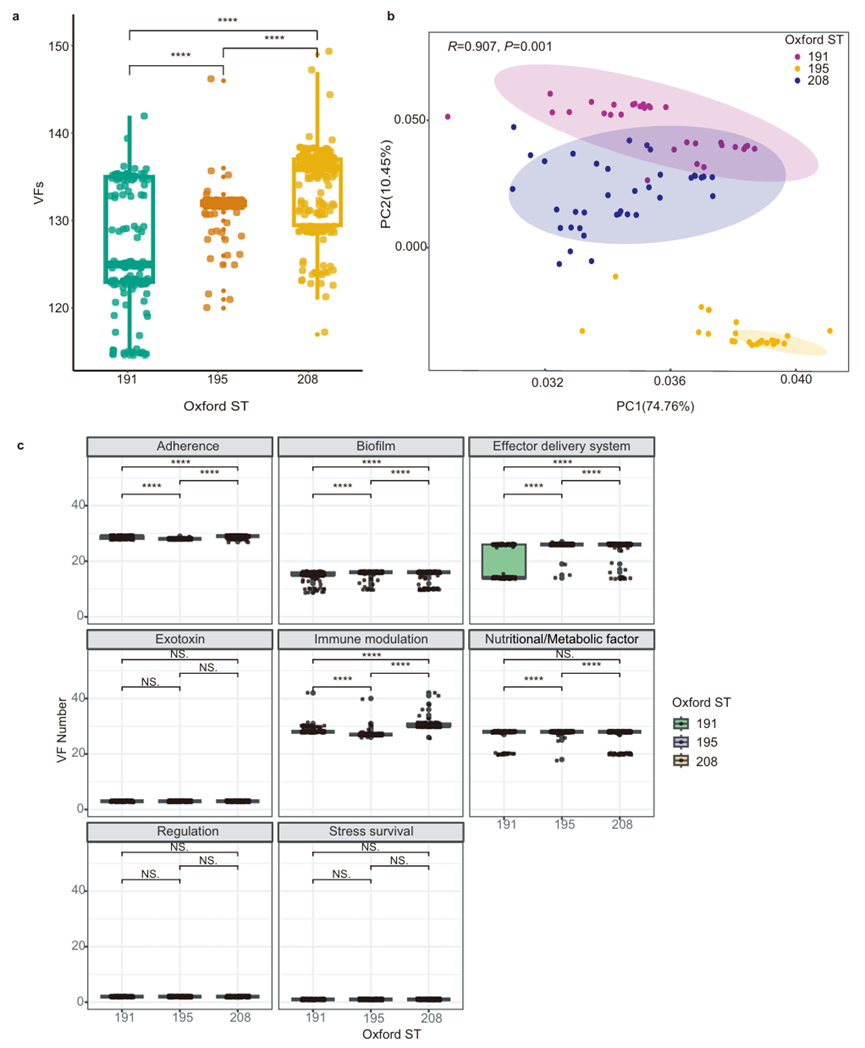
图5 ST208、ST191、ST195毒力相关基因比较。(a.毒力基因数量对比;b.毒力基因谱PCA分析;c.不同功能类别的毒力基因数量差异)
4、环境适应性和耐药性研究
①环境适应性:
在ST208、ST191、ST195菌株中共鉴定出25种消毒剂耐受基因。ST191携带数量最多,qacE基因(季铵盐抗性)在ST208和ST191中分布更多。消毒剂耐受性实验表明,ST208的消毒剂耐受表型与ST191或ST195无显著差异。
不过ST208的耐干燥性明显强于ST191和ST195,ST208-KL2表现出更高的耐干燥性,这可能为其在非生物表面的生存提供了重要优势。
②耐药性:
ARGs携带数量存在差异:IC2菌株ARGs平均携带数量(30.81个)显著多于非IC2菌株(19.59个)。
部分ARGs的携带率存在差异:IC2菌株的部分ARGs,如blaOXA-23的携带率(97.64%)显著高于非IC2菌株(11.64%)。
碳青霉烯耐药的比例存在差异:81.80%的IC2菌株为CRAB,60.36%的非IC2菌为CRAB。
ST208的ARGs数量最多:ST208菌株ARGs平均携带数量(31.97个)多于ST191(29.83个)和ST195(29.40个)。
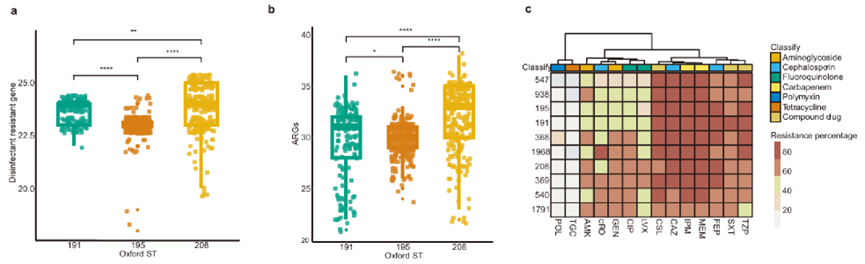
图6分离株的消毒剂耐受性和抗生素耐药性。(a.消毒剂耐受性基因数量对比;抗生素耐药性基因数量对比;c.主要ST对14种抗生素的耐药率热图)
5、基因组可塑性分析
为探究环境适应性、毒力、耐药特征差异的原因,作者进行了如下分析。
①基因组规模
ST208的基因数量(3920)显著大于ST191(3834)和ST195(3788)。
ST208的基因组长度(4.14 Mb)显著大于ST191(4.06 Mb)和ST195(4.00 Mb)。
说明ST208的基因组规模更大。
②非重组区的SNP差异
去除重组区后,ST208、ST191和ST195的核心SNP分别为2058、891和1284个。ST208的非重组区域呈现更多的SNP,特别是与细胞壁、膜、包膜生物发生、细胞外结构和碳水化合物代谢相关的基因中,可能影响环境适应性和毒力。
③ 重组热点
ST208和ST191的重组集中于荚膜多糖(CPS)和IV型分泌系统区域。
ST208-KL7的CPS区可能源自ST208-KL2,且进化分析显示ST208-KL2是所有ST208菌株的最近共同祖先(MRCA)。
ST208(32.436)和ST191(30.657)的重组率显著高于ST195(20.526)。
这些结果表明ST208获得外源基因的能力强于其它菌株,表现出更强的基因组可塑性。
④可移动遗传元件
ST208的 MGE数量和比例显著大于ST191和ST195。包括转座子(Tn)、整合性接合元件(ICE)、噬菌体(Phage)和基因组岛(Island)的数量均多于ST191和ST195。
ST208的 ICE主要携带氨基糖苷类耐药基因(AAC(6')-Ib7、APH(3')-Ia、aadA)、消毒剂耐受基因(qacE)和磺胺类耐药基因(sul1)。ST208的 Tn主要与氨基糖苷类抗性和大环内酯类抗性相关基因相关。噬菌体则没有发现耐药相关基因。这表明ST208的基因组通过ICE和Tns获得了更多的耐药基因。
为了解移动元件的差异是由垂直转移还是水平转移造成的,作者进行了MGE祖先状态重建。
⑤祖先状态重建
祖先状态重建显示,ST208的MGEs大规模扩张可能由多次获得与丢失事件引起,而非垂直遗传,表明其活跃的水平基因转移(HGT)能力。
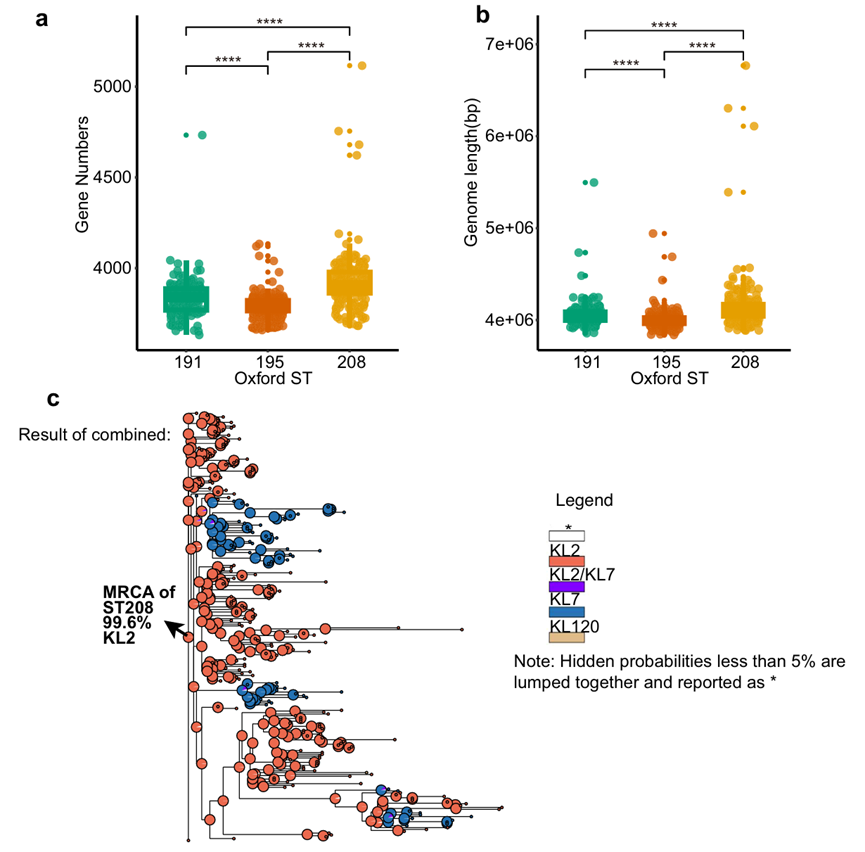
图7基因组可塑性增强促进ST208的适应性进化(a. ST191、ST195和 ST208分离株的基因数量对比;b. ST191、ST195和 ST208分离株的基因长度对比;c.基于贝叶斯二元马尔可夫链蒙特卡洛(BMM)模型构建的ST208系统发育树)
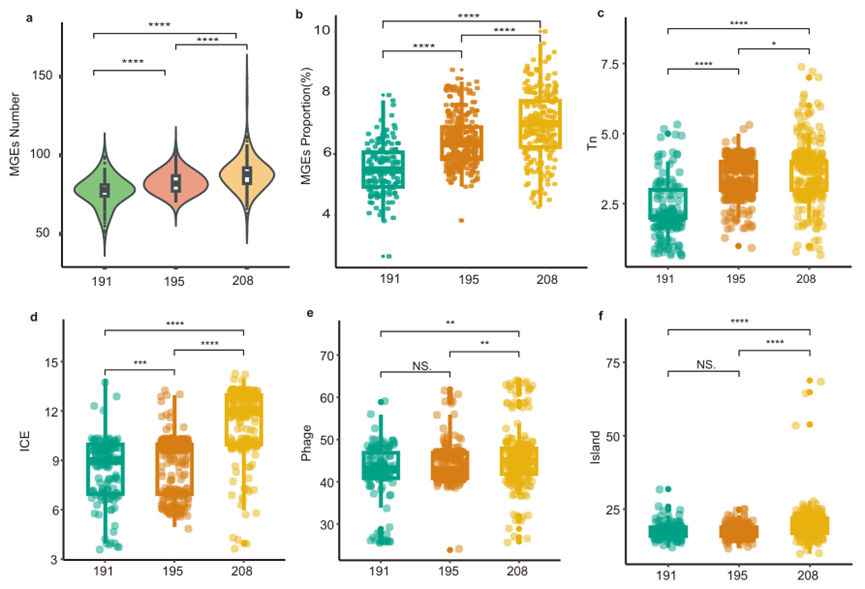
图 8 ST191、ST195和ST208分离株的可移动遗传元件(MGE)分布(a-b. MGEs的数量和比例分布;c-f. Tn、ICE、Phage、Island的数量分布)
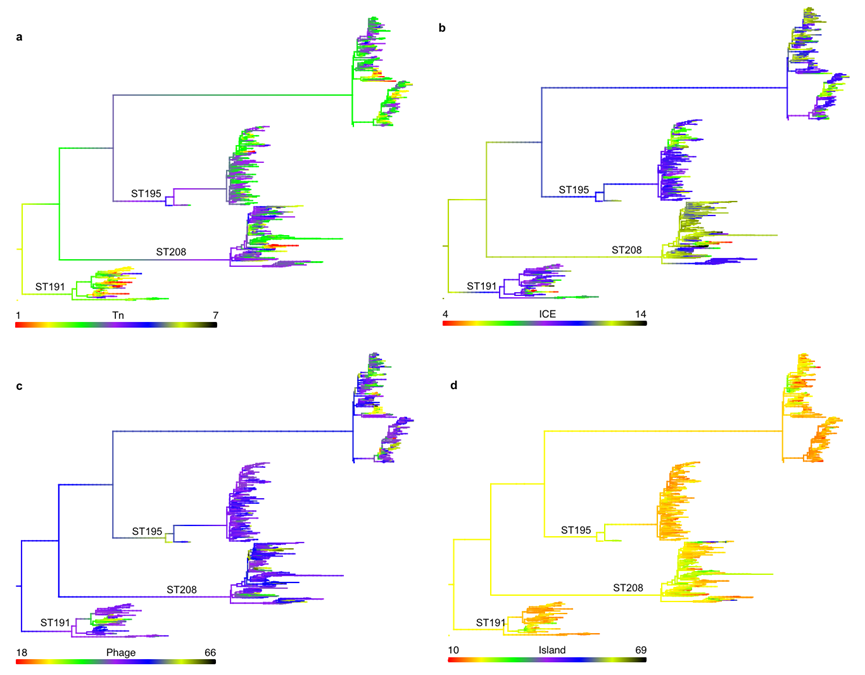
图9 MGE的祖先状态重建(MGE作为连续特征映射到基因组的系统发育树上,通过 R包 phytools重建进化,颜色代表移动元件数量)
6、系统地理学分析-传播模式
采用离散性状系统地理学推断(GTR替代模型和严格时钟模型),通过BEAST软件重建ST191、ST195和 ST208的进化历史与传播动态。
① ST191的共同祖先可以追溯到大约 1995年,起源于浙江(ZJ)。菌株按分离地点聚类,表现为克隆性医院内传播。跨省传播共26次,其中53.8%(14次)为主要传播事件,最早发生ZJ(1995年)和 AH(2001年),此后在 ZJ和 AH之间双向传播。
② ST195的共同祖先可以追溯到 1996年,同样起源于ZJ,并随后分化为两个主要分支。一个是ZJ主导分支,主要局限于本地,少量扩散至AH;另一个是全国扩散分支,祖先近期源自AH,已扩散至全国多省。跨省传播共56次,其中40次为主要传播事件,40次事件起源AH,因此AH是ST195的重要传播枢纽。
③ ST208于 1987年首次在 AH引入,随后传播到云南,中国大部分地区都能观察到不同程度的ST208流行。省际传播事件共69起,重大传播事件36起。这些传播事件涉及21个省份,AH既是最常见的传播始源地,也是目的地。
与ST191和ST195相比,ST208经历了更复杂的传输事件,传输节点更多,传播起源更早,传播范围更广,传播强度更高。值得关注的是,安徽是鲍曼不动杆菌传播的重要枢纽。
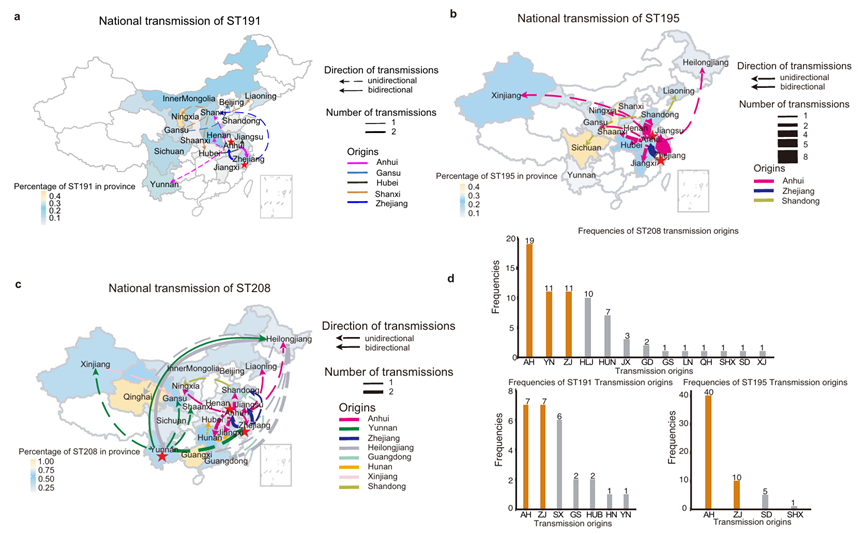
图10 ST208通过多次省际传播事件传播(a-c. ST191、ST195和 ST208的全国分布和主要传输;d.每个区域传播次数)
总结
本研究对中国76家医院2011-2021年1506株鲍曼不动杆菌血流感染分离株进行了基因组流行病学和系统动力学分析。结果显示,分离株包含149个序列型(STs)和101个K-locus类型(KLs),种群多样性增加。国际克隆(IC)2占比81.74%,其中ST208流行度上升,ST191和ST195下降,符合全球趋势。ST208相较于ST191和ST195,毒力更强、抗生素耐药性更高、耐干燥能力更强、传播模式更复杂,其基因组可塑性驱动了其适应与扩散。通过MLST (Oxford)方案,进一步揭示了ST208上升的多样性和遗传机制。研究指出,鲍曼不动杆菌正从低毒力、多重耐药病原体向高毒力方向进化,对感染控制和公共卫生政策提出迫切需求。
参考文献
Luo Q, Chang M, Lu P, et al. Genomic epidemiology and phylodynamics of Acinetobacter baumannii bloodstream isolates in China. Nat Commun. 2025;16(1):3536.
密码子云平台
我们最近推出了密码子生信云平台服务(https://cloud.mimazi.net),包含免费细菌基因组云流程和各种生信分析小工具,无需安装软件、无需配置环境,即可一键化生成数据分析及可视化绘图结果,快来试试吧!
-
点赞 (0人)
- 收藏 (0人)

