
Nat Commun.:可移动元件在碳青霉烯类抗性基因NDM全球传播中的作用
- 看不见的线
- 2948
- 2024-05-09 14:01:09
- 文章来源:bioyun

期刊介绍

背景与意义
一、研究背景
关于碳青霉烯耐药基因blaNDM的全球传播。碳青霉烯是一类广谱抗生素,由于blaNDM基因的存在,导致细菌对碳青霉烯类抗生素产生耐药性。
二、目的
本研究旨在通过对blaNDM基因周围的结构变异进行分析,重建其进化历史,并揭示其传播的主要因素。
三、意义
研究结果表明,blaNDM基因具有一个祖先背景,同时存在多个插入事件和其他基因重组,这些因素共同促使blaNDM基因在全球范围内迅速传播。
研究通过分子钟的方法,估计了blaNDM基因的起源时间,推测其在不同细菌种类中的传播途径。这项研究对了解碳青霉烯耐药性的形成和传播机制具有重要意义。
策略及方法
1.数据收集:研究人员收集了包含blaNDM基因的细菌样品的基因组测序数据。
2. 基因组组装:使用基因组测序数据,研究人员对细菌基因组进行组装,得到包含blaNDM基因的contigs(连续的DNA片段)。
3. BLAST比对:研究人员使用BLAST算法对所有含有blaNDM基因的contigs进行比对,以确定它们之间的相似性和结构变异。
4. 网络构建:根据BLAST比对的结果,研究人员构建了一个基于相似性的网络,将具有相似结构的contigs连接在一起。
5. 结构变异分析:通过分析网络中的结构变异,研究人员确定了blaNDM基因上下游的移动遗传元件,如转座子和整合子。
6. 移动元件的功能分析:研究人员对移动元件的功能进行了分析,包括它们与其他抗药基因的关联和它们在细菌中的定位。
7. 时间和地理分析:研究人员对不同时间和地理位置的blaNDM阳性菌株进行了分析,了解其传播和多样性。
为了解决研究问题,研究采用了一种基于网络的算法,将基因组序列中与blaNDM相关的序列进行分割和分类。通过这种方法,研究人员能够确定blaNDM基因的上游和下游背景,并研究这些背景中的转座子和其他可移动元件的存在和分布。此外,研究还分析了不同质粒类型之间的遗传距离和地理距离之间的相关性,以探讨质粒传播的限制因素。
总的来说,这项研究使用基因组测序数据和网络分析方法,揭示了blaNDM基因的全球传播与移动遗传元件的关系。这种方法可以为研究其他抗药基因的传播提供参考。
实验数据分析
一、blaNDM携带者的全球数据集
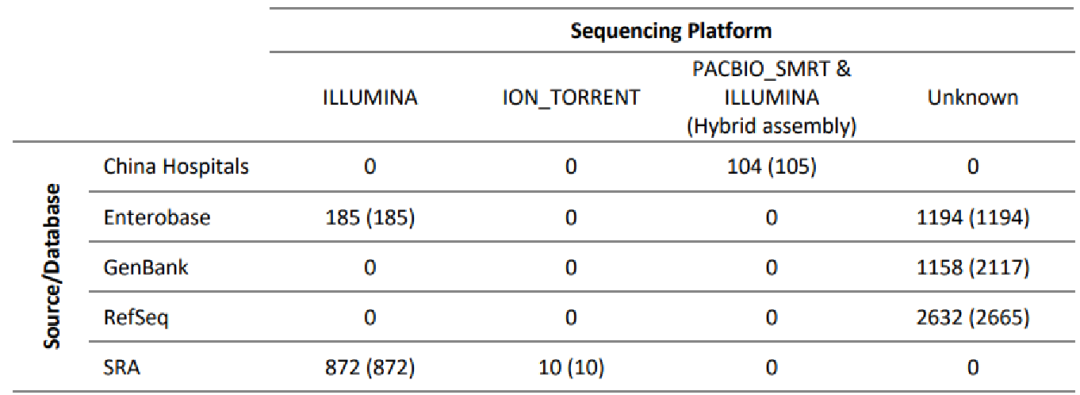
6155个细菌基因组(7148个contigs)的数据集。
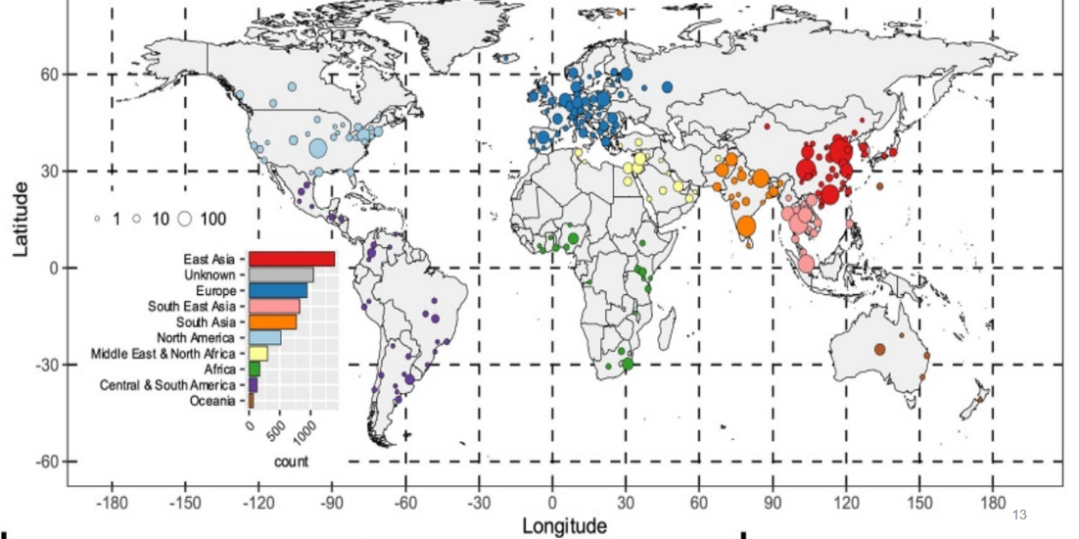
数据来自88个州,主要收集于亚洲,特别是中国(n = 1270)、欧洲国家(941)、美国(461)、泰国(419)和印度(361)。
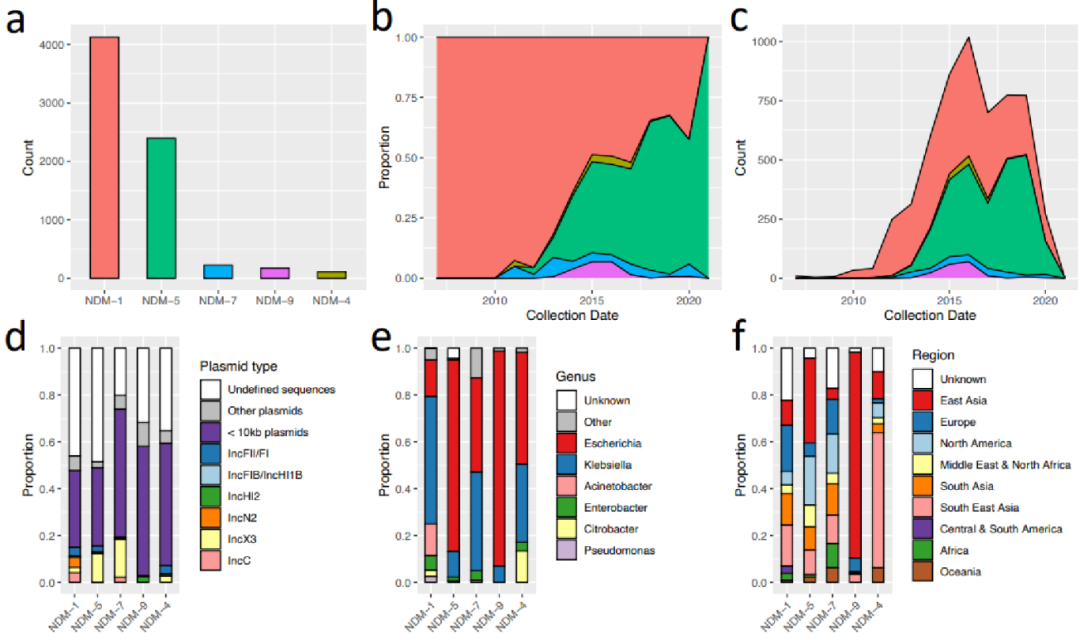
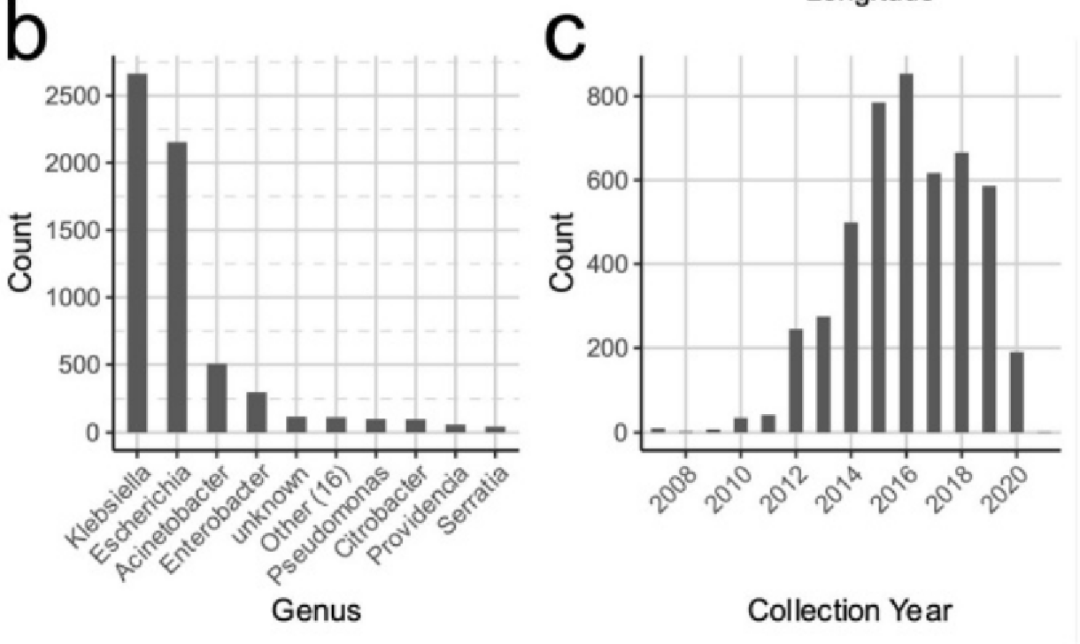
至少有27个细菌属,克雷伯氏菌和大肠杆菌分离株的比例很大(分别为2664和2154个基因组)。记录了4816份样本(78.25%)的采集日期。其中,大多数是在2014年至2019年之间收集的(71.05%)。
二、携带blaNDM的质粒类型
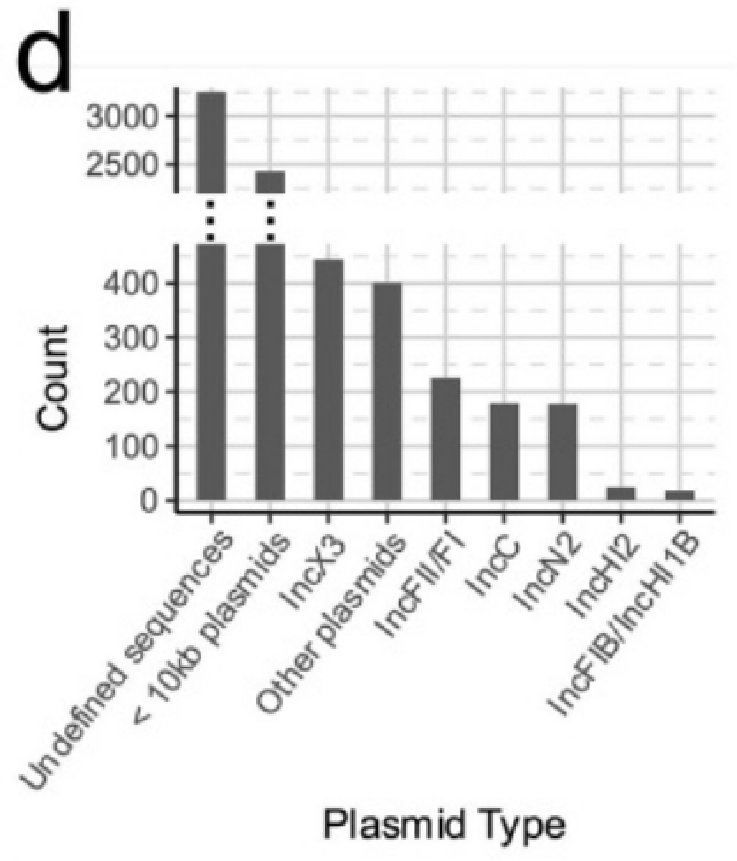
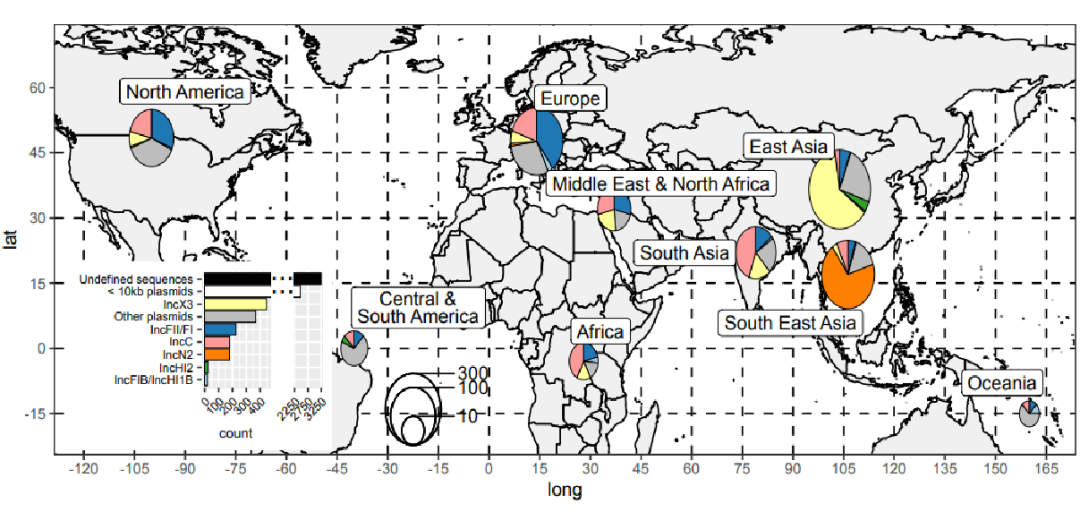
鉴定了33种不同的复制子类型。最普遍的复制子类型是IncX3(444个contigs),并且丰富的复制子类型表现出地理结构。
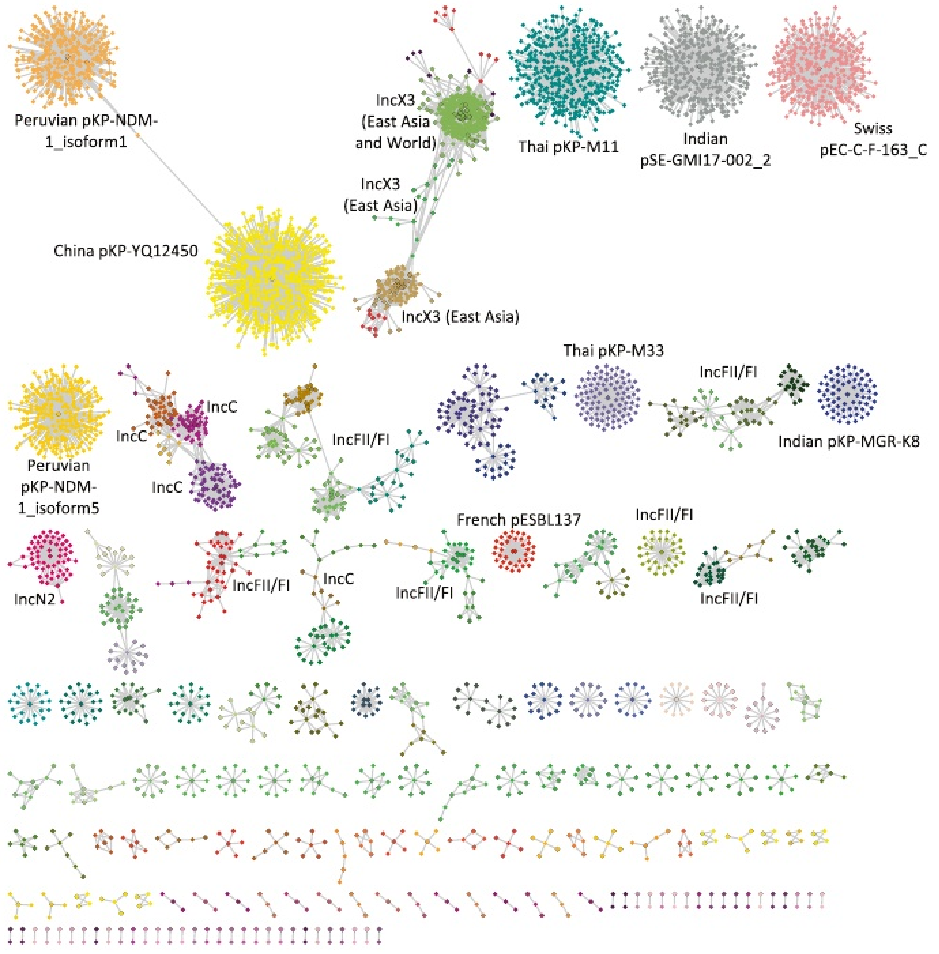
为了进一步鉴定未表征的质粒序列,将3599个contigs映射到一组完整的质粒参考序列。结果显示有181个类似的推定质粒序列簇。大多数簇(n = 105)分组的复制子类型相同,并且包含少量的contigs(只有27个簇,包含>10个contigs),这blaNDM质粒骨架的多样性和动态种群一致。
大多数(n = 2427;68.4%)的blaNDM的contigs与小型质粒(<10 Kb)相关。虽然这可能表明小质粒作为blaNDM载体发挥关键作用,但这种模式也可能是由于重复的ISs和转座子导致的持续碎片化的从头组装造成的。
三、解析blaNDM侧翼区域的结构变异
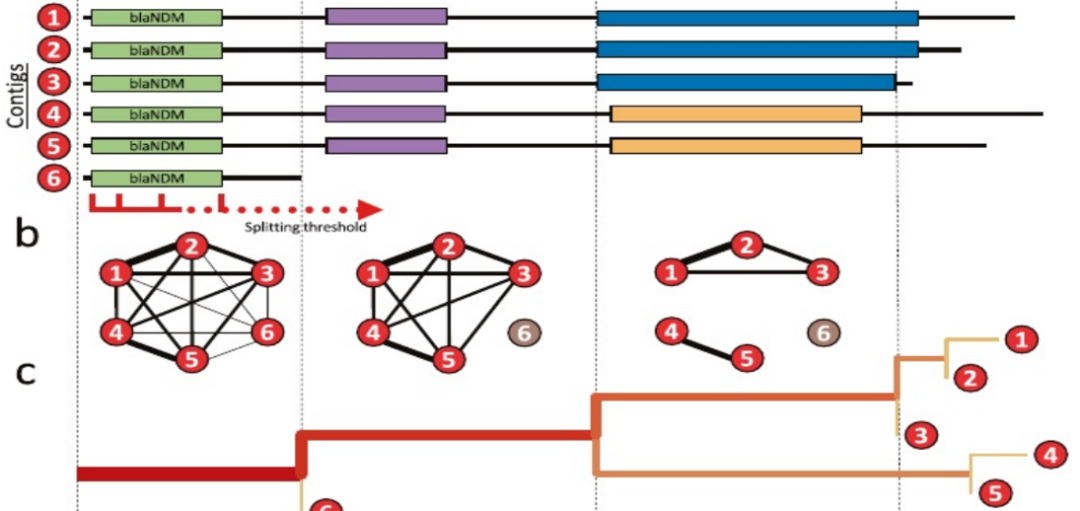
对所有NDM阳性的Contigs进行配对BLAST搜索。从blaNDM开始,继续向下或向上,使用“分裂阈值”逐渐增加被检查区域。在每个步骤中,构建一个图,连接具有“分裂阈值”给定的最小长度的共享BLAST命中的contigs(节点)。在阈值的某一位置具有相同结构变体的Contigs属于同一图分量,而短的Contigs被挑出来。分裂被可视化为一个树,其中分支长度被缩放以匹配序列内的位置,分支的厚度和颜色强度对应于携带同源性的序列的数量。
四、传播blaNDM的早期事件
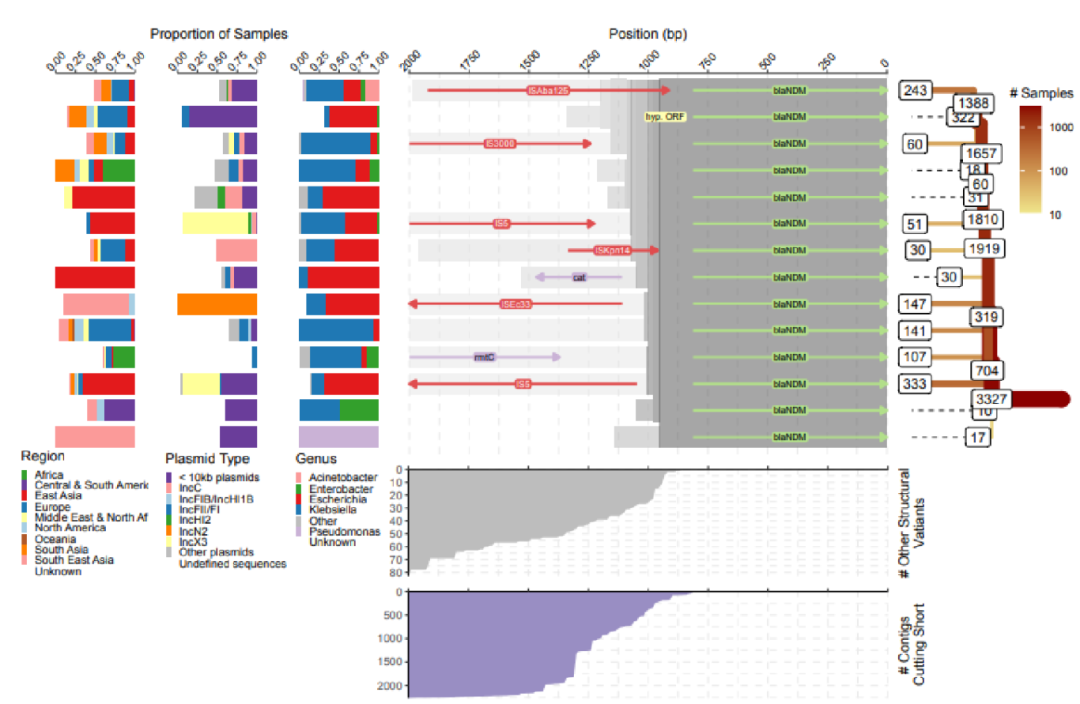
blaNDM上游的结构变异
表明Tn125是blaNDM基因的祖先转座子。blaNDM上游区域的同源性迅速下降:在blaNDM起始密码子的几百个碱基对内,该区域分裂成多个结构变体,没有一个在已有contigs库中占主导地位。
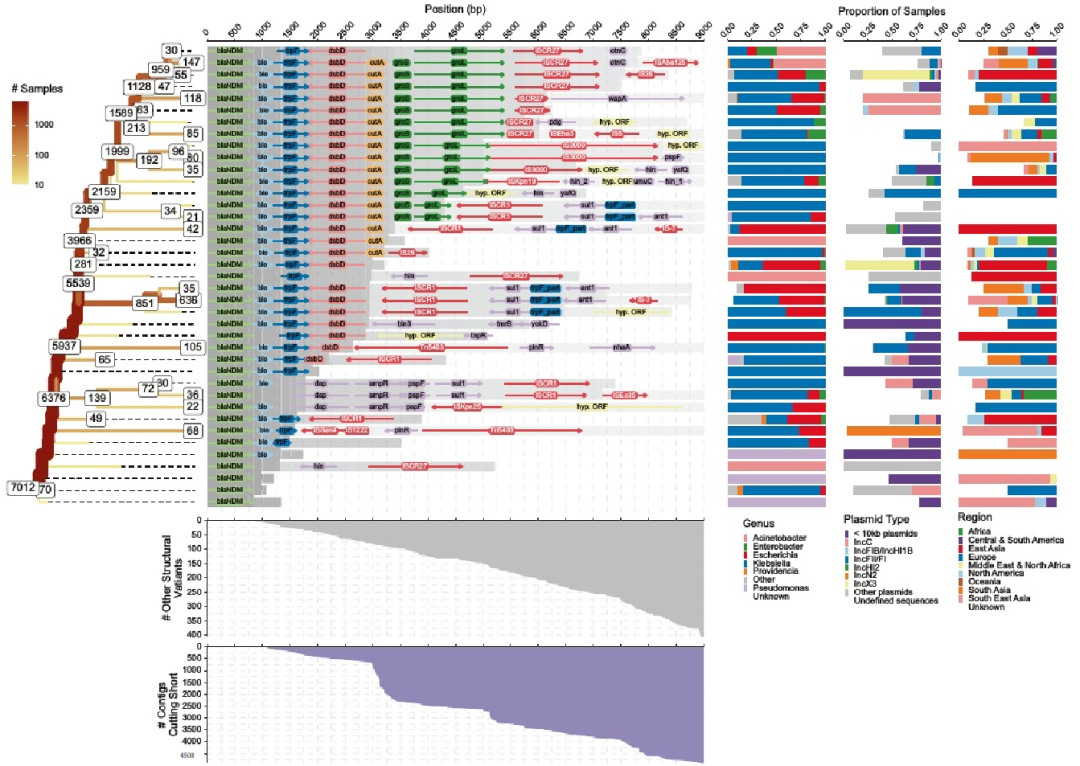
下游侧翼区域比上游区域表现出更渐进的结构多样化,具有一个主要的假定祖先背景。从下游图中得知,假定的祖先Tn125背景,具有不间断的下游ISAba125元件,主要来自不动杆菌和克雷伯氏菌。相反,上游携带ISAba125的细菌属的多样性更为丰富。这支持了Tn125最初将blaNDM传播到由不动杆菌和克雷伯氏菌主要介导的其他质粒上,之后转座子被其他重排破坏。
IS3000,无论是上游还是下游,几乎都只与克雷伯氏菌的样本相关。Tn3000由两个IS3000拷贝组成的复合转座子——可能在Tn125“石化” 后重新激活blaNDM,这种二次动员主要发生在克雷伯氏菌物种中。Tn5403被发现与IncN2质粒广泛相关,这可能通过协整中间体将blaNDM置于这种背景中。
下游侧翼区最常见的转座因子之一是ISCR1 (IS91家族转座酶),它总是伴随着sul1,偶尔与ant1或ppf、ampR和dap基因配置。ISCR1的取向应该阻止该元素移动blaNDM,这种元素的流行可能是由于它可以调动几个抗菌素耐药性基因。ISCR27,它一直在groL基因的下游被发现,出现率很高,正确定向动员blaNDM,这被认为是在blaNDM祖细胞的初始动员过程中发生的。
五、随后的重排以IS26为主
在blaNDM下游的特定距离(例如,区域3000-3300 bp),考虑的contigs数量的三次急剧下降促使我们研究这些不同的截断点。
与观测到的最大落差对应的blaNDM下游3000-3300长侧翼相关的≥50 bp的overhanging reads几乎完全匹配IS26序列的左侧倒置重复。
与IS26相关的另一个热点在7500 bp左右被发现,而在7800 bp左右,许多overhanging reads映射到ISAba125。这些位置大致与blaNDM下游7500-8000 bp处观测到的contigs数量的第三个下降相匹配。
IS26虽然经常在两个相邻的拷贝中发现,形成一个看似复合的转座子,但它是一个所谓的伪复合转座子。与复合转座子相反,两个IS26侧翼的DNA片段通过共整形成或以圆形可转位单元(TU)的形式被动员,其由单个IS26元件和被动员的DNA片段组成,并优先插入另一个IS26旁边。
六、进一步研究blaNDM的移动性
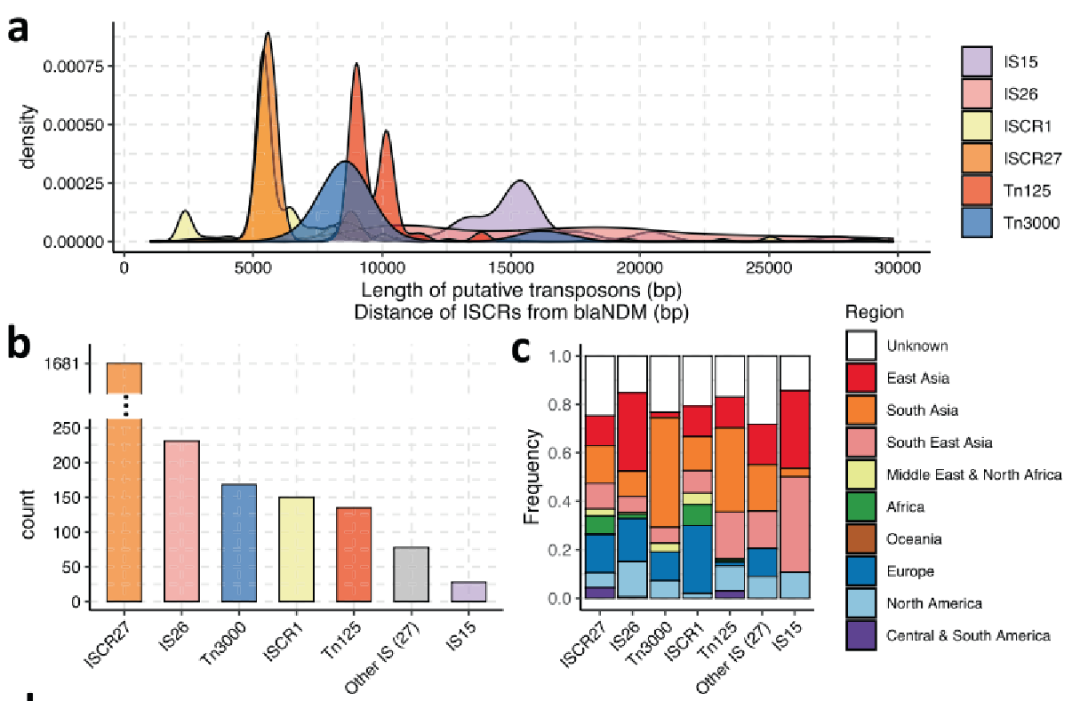
对理论上能够动员blaNDM的最常见(伪)复合转座子进行了表征。这些转座子被定义为DNA的延伸,两侧有两个匹配的完整或部分ISs,间隔<30 Kb,并包围blaNDM。
总共在468个contigs中鉴定出640个复合转座子,包括31种不同类型,其中最常见的是:IS26(231个),IS3000(形成Tn3000;168个),ISAba125(形成Tn125;138个),IS15(28个)。Tn125和Tn3000复合转座子的长度在7-10 Kb之间是一致的。
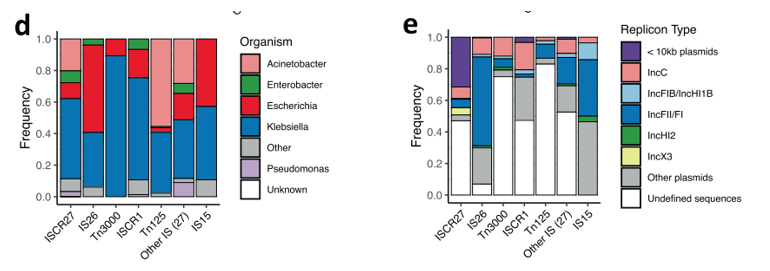
在东亚和东南亚收集的样本中,IS15和IS26的存在增加了。这些在大肠杆菌菌属和克雷伯氏菌属中大致相同地发生,并且与多个质粒主干相关,但主要发生在IncF质粒上。Tn125和Tn3000在印度次大陆具有显著的优势,并且分别主要存在于不动杆菌属和克雷伯氏菌属中。
七、关键事件的分子钟
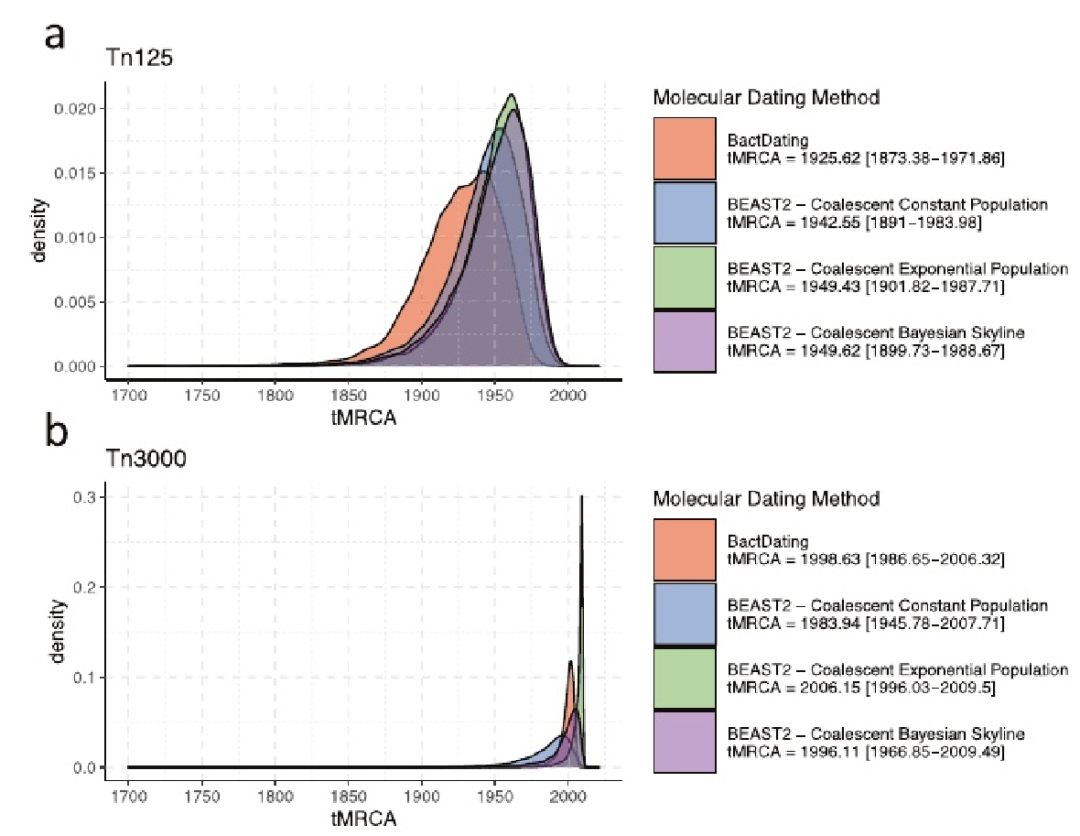
估计了Tn125和Tn3000转座子形成的相对时间,与Tn125相比,我们在Tn3000序列中观察到更少的SNP,主要位于转座酶基因内贝叶斯分析表明,携带blaNDM基因的Tn125转座子最近的共同祖先(MRCA)可以追溯到1990年之前。Tn3000对blaNDM的动员估计发生在千禧年之交的晚些时候。这些发现与更广泛的说法是一致的,即blaNDM的传播最初是由Tn125动员驱动的,随后由Tn3000、IS26和其他因素转位。
八、blaNDM分离株的时间多样性提示了质粒的作用
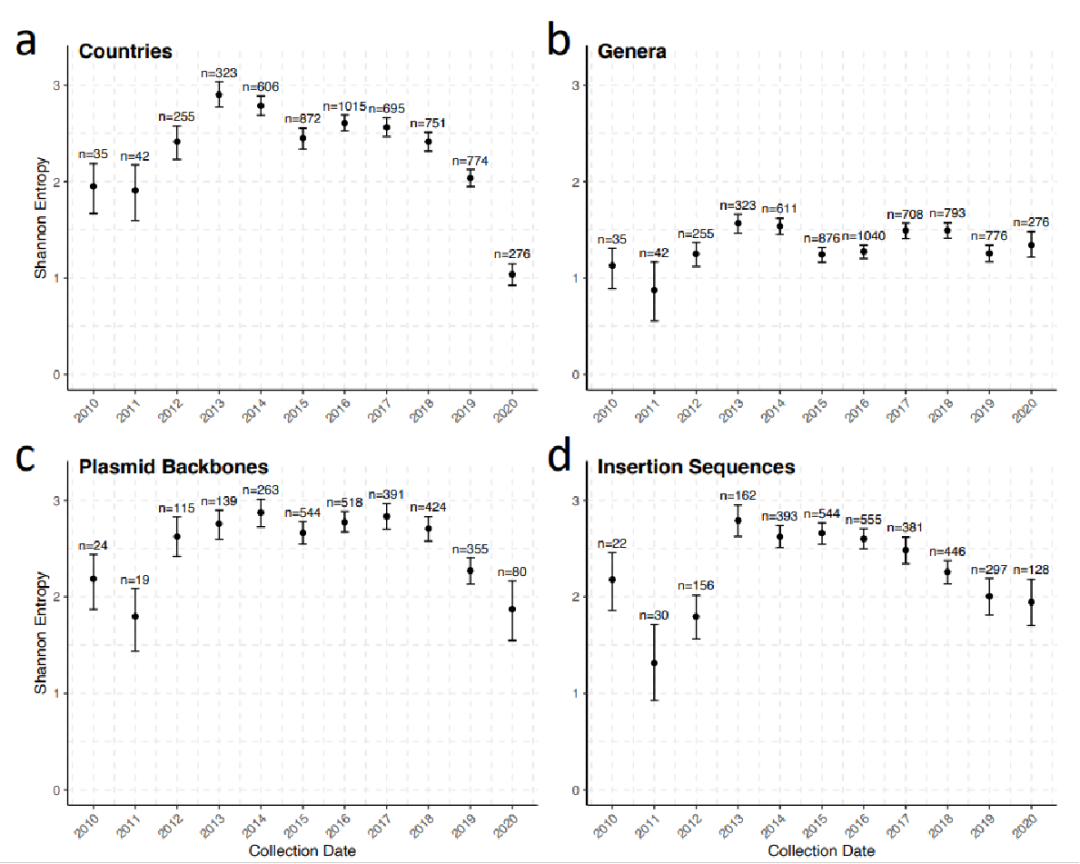
图中较高的熵值表示具有丰富的等可能的类别,而较低的熵值表示有限数量的类别,用blaNDM分离株相关的国家多样性的变化来近似blaNDM全球传播的广泛模式。NDM的全球传播持续了8-10年。
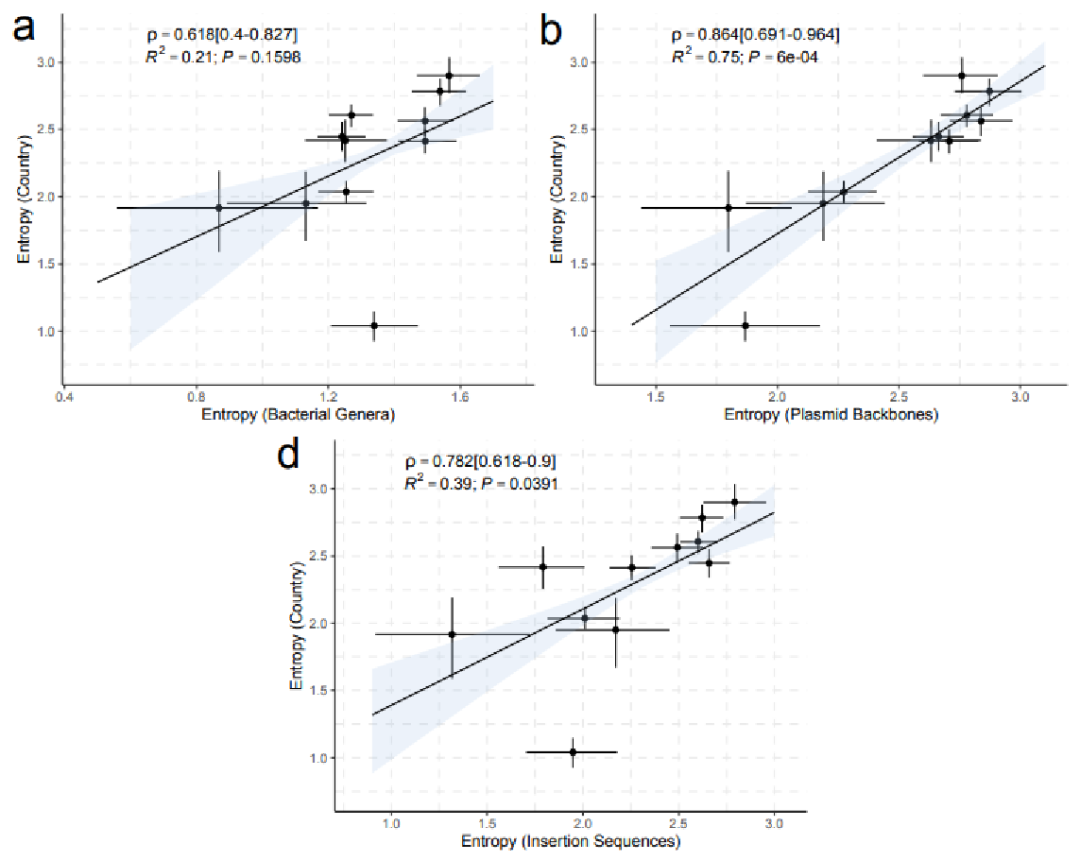
与blaNDM分离菌相关的国家多样性的估计变化与其他元数据类别呈正相关。在国家多样性和质粒主干之间发现了最强的相关性。
没有检测到IncX3和IncN2质粒之间的关系,因为这两种复制子类型大多分别局限于中国和印度。在IncF和IncC以及其他已确认的质粒序列中,随着更多的质粒序列被纳入分析,在下游发现了显著的逐渐增加的相关性强度。
这些趋势表明,携带blaNDM的质粒具有地理结构,而blaNDM的传播基本上是一个空间过程。这将与质粒生态位的存在相一致:即特定质粒更能适应的环境。
结果与讨论
blaNDM基因的全球传播主要受到移动基因元件的影响,其中IS26-包围的伪复合转座子是blaNDM基因在近期遗传重组中最重要的贡献者。此外,研究还发现了多种携带blaNDM的质粒类型,并且发现了质粒骨架长度的遗传距离与采样地点的地理距离之间存在正相关关系。
理论意义在于揭示了blaNDM基因的全球传播机制,并提供了对移动基因元件在抗生素耐药性传播中的作用的深入理解。实践意义在于为控制和预防抗生素耐药性的传播提供了重要的参考,为制定合理的抗生素使用策略和防控措施提供了科学依据。
总结与启发
一、创新性
采用网络算法来重构blaNDM基因的上下游背景,并通过分析移动基因元件的角色揭示了blaNDM基因的全球传播机制。此外,研究还发现了IS26包围的伪复合转座子在blaNDM基因的遗传重组中起到重要作用。
二、不足
仅针对blaNDM基因进行了分析,对其他移动基因元件的传播机制尚未进行深入研究。此外,由于方法的局限性,上下游背景的重构仅限于blaNDM基因,对于其他移动基因元件的重构仍然存在困难。
三、展望
根据本研究的结果,后续可能的研究方向包括对其他移动基因元件的传播机制进行深入研究,以及进一步探索移动基因元件与抗生素耐药性传播之间的关系。此外,可以考虑开发更高效的方法来同时重构基因的上下游背景,以便更全面地了解移动基因元件的遗传重组过程。
原文链接:
https://www.nature.com/articles/s41467-022-28819-2
-
点赞 (0人)
- 收藏 (0人)

