
【文献速递】高毒肺炎克雷伯菌如何杀伤肠道竞争菌?
- 看不见的线
- 4302
- 2024-06-11 08:52:58
- 文章来源:临床与微生物

【据《Isme Journal》2024年3月报道】题:Hypervirulent Klebsiella pneumoniae employs genomic island encoded toxins against bacterial competitors in the gut(作者:Yi Han Tan等)。
【摘要】
高毒力肺炎克雷伯菌 (HvKp)会导致侵袭性感染,例如克雷伯菌肝脓肿。侵袭性感染通常发生在 HvKp 最初定植宿主胃肠道后。超过 80% 的 HvKp 分离株属于克隆组23亚系I(CG23-I),该亚系已获得基因组岛 (GI) GIE492 和 ICEKp10。本研究分析了 12361 个肺炎克雷伯菌基因组,现基因毒力岛GIE492和ICEKp10与CG23-I和CG10118的HvKp谱系共同相关;③GIE492和ICEKp10编码的细菌素microcin E492(mccE492)和基因毒素colibactin增强了HvKp在肠道的定植能力,其中colibactin主导了菌群多样性变化;体外实验表明,colibactin和mccE492能够杀死或抑制多种革兰氏阴性菌如克雷伯菌和大肠杆菌等,以及多种革兰氏阳性细菌如双歧杆菌和毛螺菌属等;研究结果表明,mccE492破坏细菌细胞膜,使基因毒素colibactin更好地进入胞内实现基因毒性,两种毒力岛的获得可能有助于HvKp在宿主中更好地定植,解释了CG23-I谱系的优势地位。
【引言】
肺炎克雷伯菌是一种革兰氏阴性菌,是一种与肺炎相关的院内病原体。高毒力肺炎克雷伯菌(HvKp)是社区获得性的,是亚洲单菌性肝脓肿的主要原因。HvKp的优势谱系是CG23-I,主要由序列23型(ST23)菌株组成,这些菌株导致约80%的克雷伯菌肝脓肿(KLA)感染。在亚洲国家,HvKp的胃肠道携带率可能很高,在新加坡的一项研究中,腹泻患者的胃肠道携带率为3%至8%,在韩国的一项研究中,健康人的胃肠道携带率为21.1%。ST23 HvKp仅从患者而非环境样本中分离出来,肠道定植与KLA的发展密切相关。因此,有利于HvKp持续胃肠道携带的因素可能有利于其优势和传播。
基因组岛(GIs)是整合在细菌染色体中的水平获得的DNA元件。GI有助于细菌物种的辅助基因组,并且可以包含多种遗传物质,例如代谢操纵子和毒力决定因子。大多数肺炎克雷伯菌染色体至少有4个GIs,有些菌株拥有多达10个GIs,许多编码已知或假定的毒力因子。由于GI可以赋予有益的表型,它们推动了物种内某些细菌谱系的快速进化和成功。大多数HvKp菌株具有一个编码毒力因子的大毒力质粒,以及几个GIs。GIE492和整合共轭元件Kp10 (ICEKp10)与CG23-I谱系密切相关。目前缺乏关于肺炎克雷伯菌群体中GIE492的进化历史和流行情况的研究。此外,GIE492和ICEKp10在人类宿主HvKp发病机制中的作用尚不清楚。
GIE492含有合成和输出细菌素 microcin E492 (mccE492)所需的mce基因位点。
MccE492 是一种铁载体微菌素,可通过特洛伊木马机制杀死其他细菌。MccE492 是一种铁载体微菌素,通过外膜上的儿茶酚铁载体受体进入易感细胞的周质 。随后,mccE492 插入内膜导致细胞死亡。ICEKp10 携带编码耶尔森菌素铁载体的 ybt 基因座,而 clb 基因座能够产生colibactin。Colibactin 是一种小分子烷基化基因毒素,可导致双链 DNA 断裂。肠道携带 clb 阳性细菌与结直肠癌相关。最近的研究表明,除了对宿主的影响之外,产colibactin的大肠杆菌还可以利用colibactin来杀死其他细菌。
我们分析了 GIE492 在肺炎克雷伯菌中的进化和流行情况,发现了 GIE492 的八个主要变体,其中一个特定变体与高毒力 CG23-I 肺炎克雷伯菌谱系中的 ICEKp10 共同相关。此外,我们发现GIE492和ICEKp10在SGH10(CG23-I的代表性临床分离株)胃肠道定植过程中发挥协同作用。Colibactin 会导致肠道微生物发生变化,有利于 HvKp 定植。我们的结果支持GIE492 和 ICEKp10 使 HvKp 能够与肠道生态位中的其他细菌物种有效竞争。因此,我们假设 GIE492 和 ICEKp10 的获得可能是CG23-I 成为 HvKp 的主要谱系的一个促成因素。
【结果】
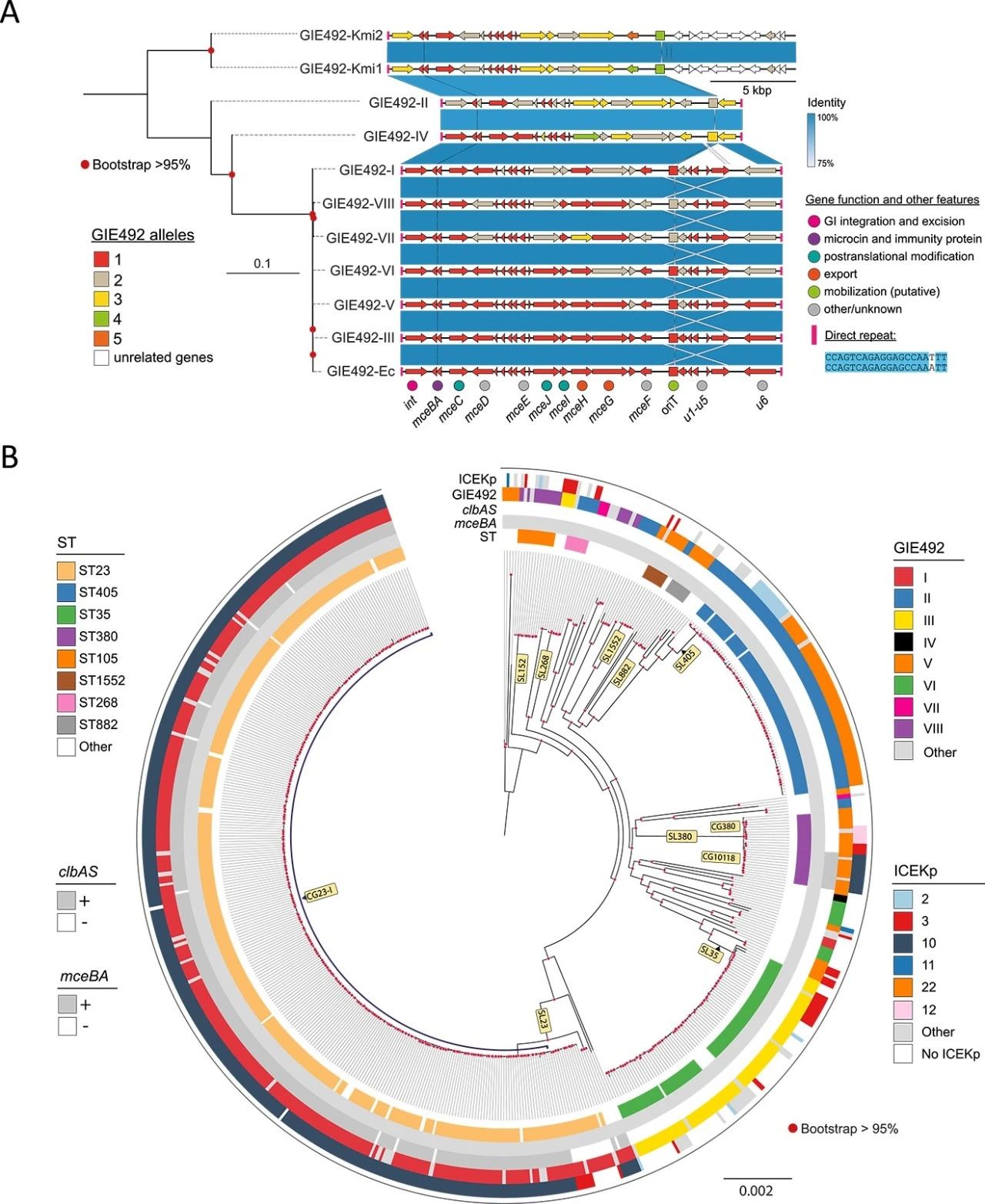
1.GIE492 的进化史及其与肺炎克雷伯菌 CG23 中 ICEKp10 的关联
我们在 12433 个 KpSC 基因组中搜索了 GIE492,从中鉴定了 657 个 GIE492+ 基因组,并根据序列比较和对GIE492中23个基因的等位基因发现了 8 个主要的 GIE492 变体。六个变体的长度与 GIE492-III 相似(首先在 RYC492 中表征),并且 GIE492-II 和 GIE492-IV 缺乏包含功能未知的 u1-u5 基因的约4.6kbp 区域(图 1A)。
我们还在 RefSeq 数据库的一组 82206 个肠杆菌基因组中搜索了 GIE492。除肺炎克雷伯菌外,GIE492仅在两种大肠杆菌(GIE492-Ec)和两种密歇根克雷伯氏菌基因组(GIE492-Kmi1和2)中发现(图1A)。后者携带与GIE492-II更相似的不同GIE492,在3’区域有重排和不同的基因内容,缺乏直接重复(图1A)。因此,GIE492 高度局限于肺炎克雷伯菌,尽管它可能以相对较低的频率转移到其他相关物种。
为了研究 GIE492+ 菌株之间的系统发育关系,使用 cgMLST 分析鉴定了亚谱系和克隆组。SL23、SL405、SL35(包括RYC492)、SL380和SL152最为常见,并且分别主要与变体I、II、III、V和VIII相关(图1B)。这种分布表明该 GI 在不同的肺炎克雷伯菌谱系中早期获得,随后在每个谱系内垂直传播。
很大比例 (82%) 的 GIE492+ 基因组携带 ICEKp 元件。GIE492-I 和 ICEKp10 在全球传播的 HvKp 亚支 CG23-I 中同时出现(图 1B)。CG23-I与等位基因ybt1、clb2和毒力质粒KpVP-1高度相关。此外,GIE492-V和ICEKp10也在CG10118(SL380)中同时出现(图1B)。特定 GIE492 变体和 ICEKp10表明,GIE492 发生了两次独立的获得,并保存在这些高毒力进化枝中。CG23-I 是 HvKp 的主要谱系,GIE492 和 ICEKp10 可能对其成功做出了贡献。
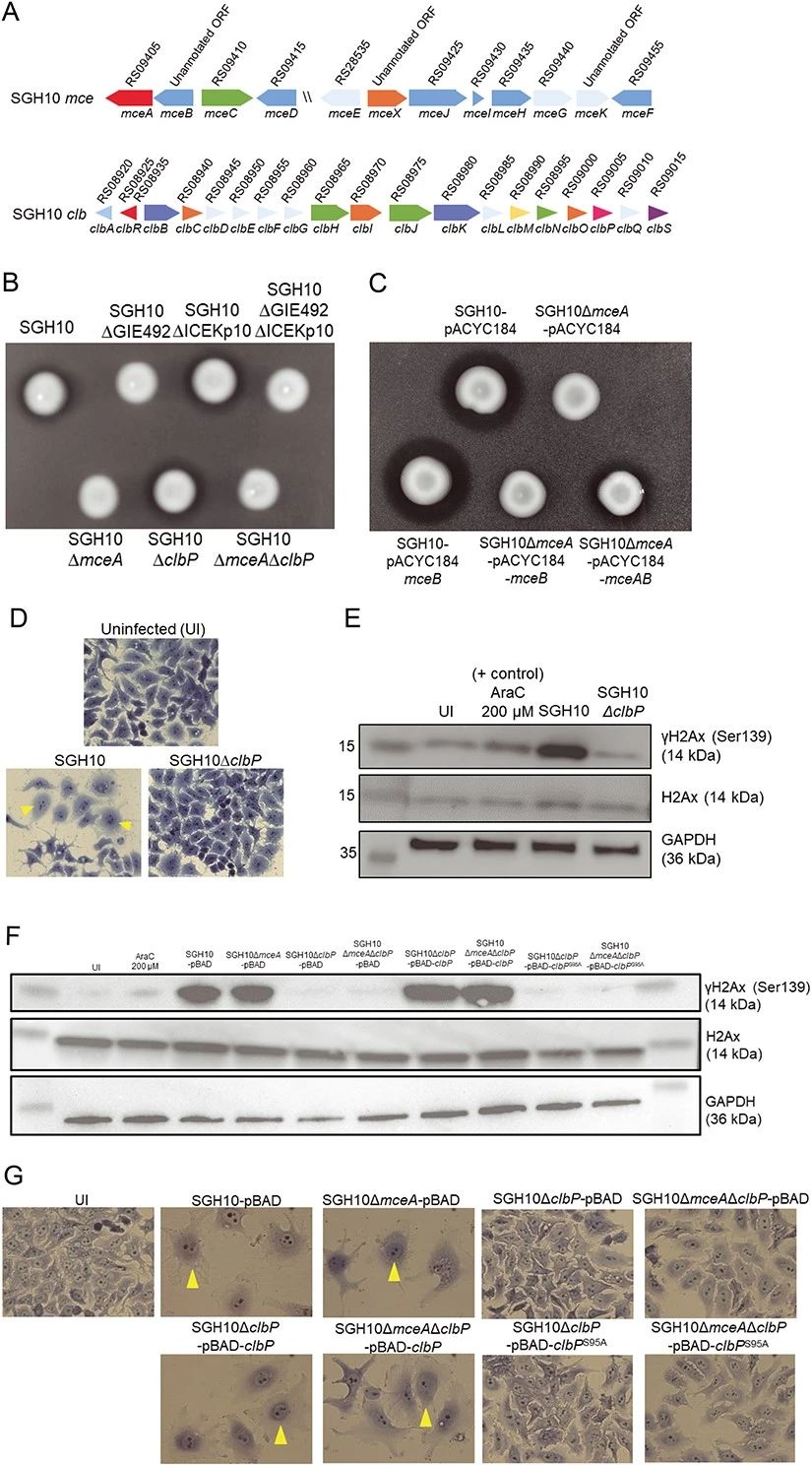
2.SGH10 产生功能性 mccE492 和colibactin
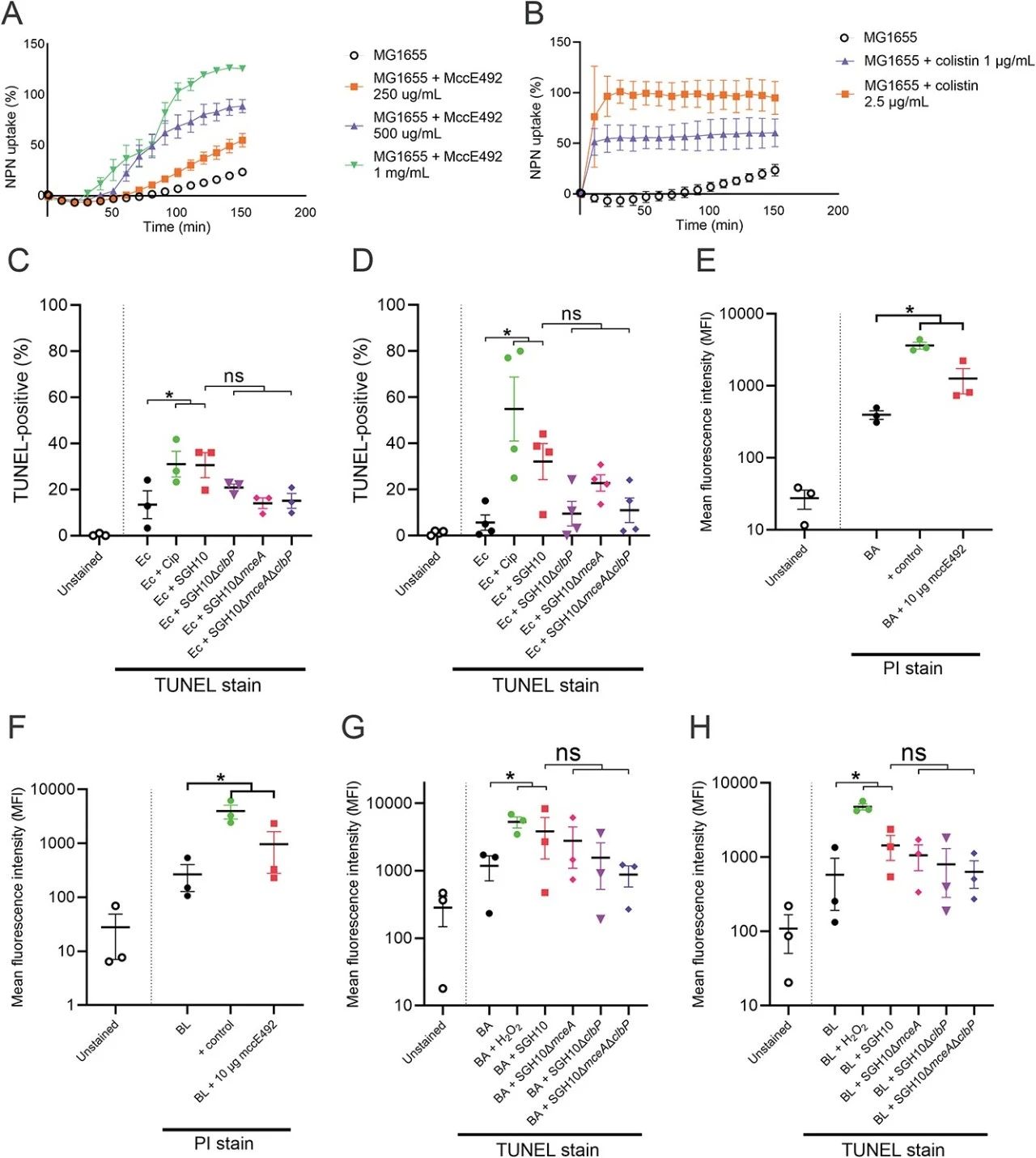
SGH10 中的 mce 和 clb 基因位点分别编码小菌素 E492 和colibactin(图 2A)。我们创建了 GIE492、ICEKp10、mceA 和 clbP 的基因缺失突变体。使用琼脂扩散测定证明 SGH10 产生功能性 mccE492,抑制大肠杆菌猎物的生长(图 2B)。GIE492 和 mceA中的缺失突变体不会杀死大肠杆菌(图 2B)。由于单独表达过量的 mceA 会导致自中毒,因此我们通过在天然启动子的控制下表达mceAB来补充 SGH10ΔmceA(图 2C)。互补菌株杀死了大肠杆菌猎物,而单独表达 mceB 的 SGH10ΔmceA 或空载体却没有(图 2C)。为了验证 SGH10 产生colibactin的能力,HepG2 细胞被 SGH10 感染。受感染的细胞表现出扩张的细胞核和细胞体,这是由colibactin的基因毒性作用诱导的巨细胞表型的特征(图2D)。相反,用ΔclbP感染的细胞(其中负责colibactin成熟的酶已被敲除)在形态上与未感染的细胞相似(图2D)。γH2AX 积累是哺乳动物细胞中双链 DNA 断裂的标志物。我们在感染 SGH10 的 HepG2 细胞中观察到强烈的 γH2AX 信号,但在感染 SGH10ΔclbP 的细胞中观察不到(图 2E)。SGH10ΔclbP 与野生型 clbP 的互补恢复了遗传毒性(图 2F)。ClbPS95A(一种催化失活的 ClbP)的表达不会恢复 SGH10ΔclbP 中的γH2AX 积累或巨细胞作用(图 2F 和 G)。
3.MccE492和colibactin对肠道定植很重要
由于 GIE492 和 ICEKp10 与 CG23-I 谱系共同关联,我们假设这些 GI 在播散性感染过程中很重要。在全身感染的小鼠模型中,SGH10ΔGIE492和SGH10ΔICEKp10感染肝脏和脾脏的能力没有缺陷,尽管肺部的SGH10ΔICEKp10略有减少(图3A)。第11天和第13天,SGH10ΔICEKp10的粪便CFU显着低于SGH10(图3B)。SGH10ΔGIE492和SGH10的粪便CFU没有差异(图3B)。因此,ICEKp10 而不是 GIE492 对于胃肠道持久性很重要。ICEKp10和GIE492除了mce和clb之外还含有其他遗传内容。然后我们创建了 ΔmceA 和 ΔclbP 来确定 mccE492 和 colibactin 的具体作用。SGH10ΔmceA 以及 SGH10 均定植(图 3C)。然而,从第11天到第17天,SGH10ΔclbP和SGH10ΔmceAΔclbP在肠道中持续存在的能力减弱(图3C)。第13天和第15天,SGH10ΔmceAΔclbP的粪便细菌载量显着低于SGH10ΔclbP(图3C)。肠道定植似乎是由colibactin驱动的,而 mccE492 通过放大colibactin的作用发挥支持作用。
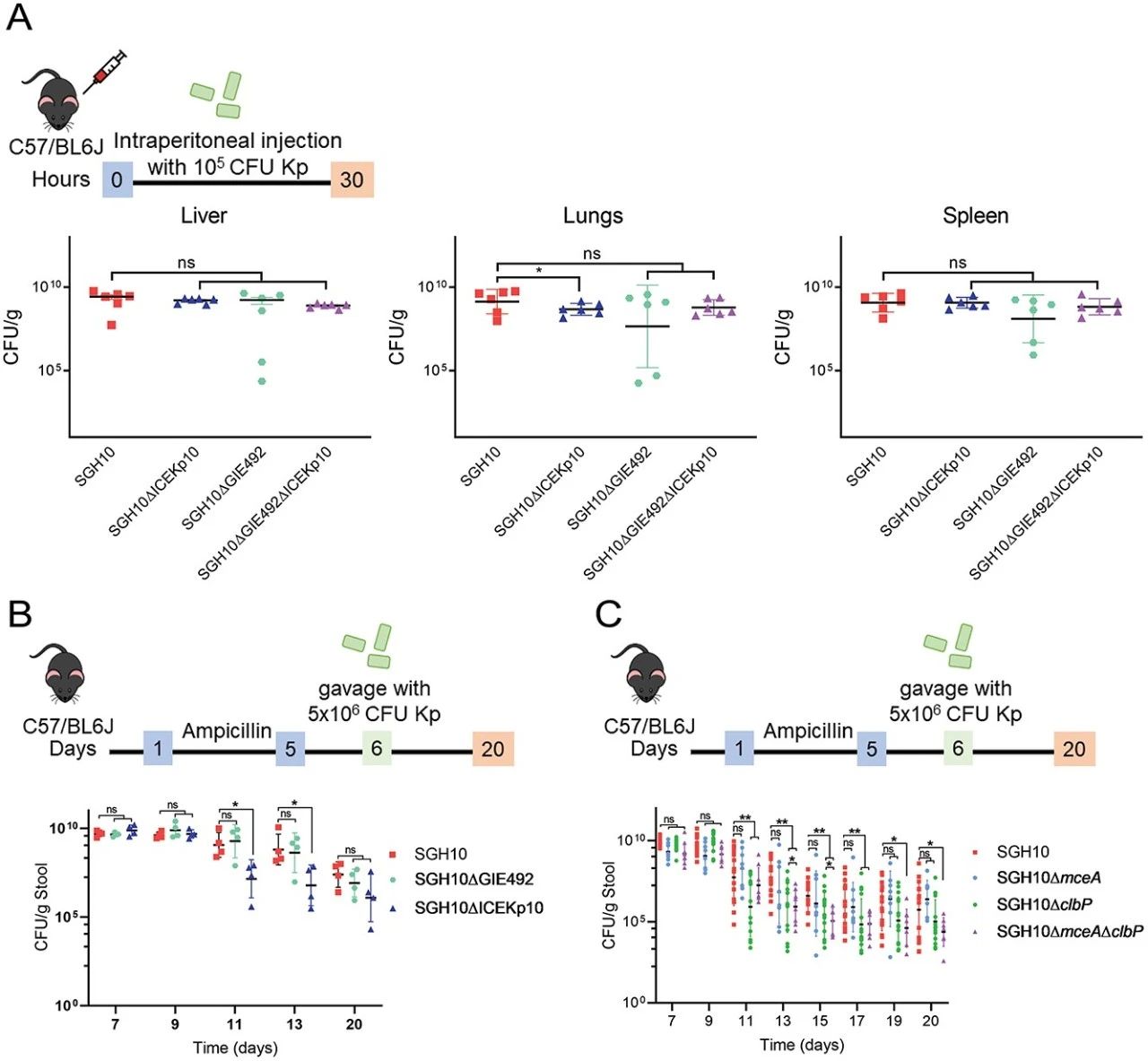
我们开发了一种依赖于抗生素混合物和 Miralax 的肺炎克雷伯菌易位小鼠模型,Miralax 是一种温和的渗透性泻药,可诱导肠粘膜变薄。然后我们确定colibactin在胃肠道易位的情况下是否重要(图4A)。大约85.7%感染 SGH10 的小鼠死于感染,而 SGH10ΔclbP 仅杀死 35.7% 的感染小鼠(图 4B)。第 8 天粪便细菌载量存在显着差异(图 4C),表明该模型中 SGH10ΔclbP 的减弱可能是由于肠道定植的短暂差异所致。colibactin也可能会导致肠上皮损伤并增加 SGH10 的易位。

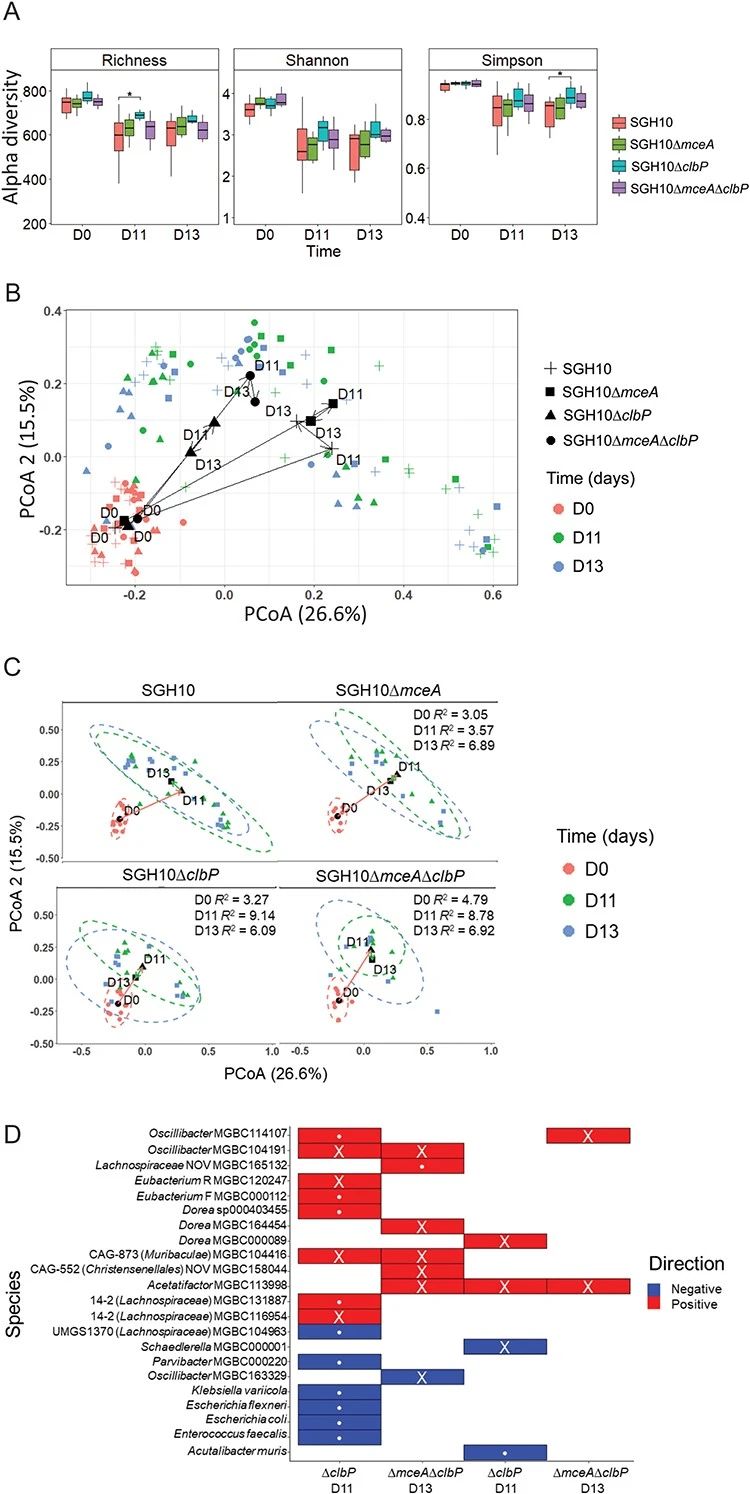
4.colibactin驱动肠道微生物组变化
为了确定肠道细菌分类群是否受到影响,对图 3C 中的实验样本进行了鸟枪法宏基因组测序。在用氨苄青霉素干扰微生物群之前(第 0 天),以及在组间观察到肺炎克雷伯菌丰度最大差异时(第 11 和 13 天),收集粪便。我们在第 11 天和第 13 天观察到肠道微生物组中与colibactin相关的 α 多样性减少,但与其他突变体无关(图 5A)。随后,我们在 PCoA 图上绘制了Bray-Curtis 距离(图 5B)。第 0 天的点聚集得更紧密(图 5B),但在氨苄青霉素处理和肺炎克雷伯菌定植后,根据细菌突变体的不同,出现了不同的变化(图 5C)。对不同组样本之间的 Bray-Curtis 距离进行置换多元方差分析 (PERMANOVA),结果表明SGH10 与 SGH10ΔclbP 的肠道微生物组组成在第 11 天和第 13 天均存在显着差异,以及 SGH10 和 SGH10ΔmceAΔclbP 感染小鼠之间,但仅在第 11 天(图 5C)。与 SGH10 相比,我们没有观察到感染 SGH10ΔmceA 后微生物组组成存在显着差异,并且观察到 SGH10ΔmceA 的 R2 值较小(图 5C),表明肠道微生物组变化的主要驱动因素是大肠杆菌素。
5.肺炎克雷伯菌通过microcin和colibactin依赖性机制杀死其他细菌
虽然肠道定植不需要 mccE492,但 HvKp 在肠道中同时表达 mccE492 和colibactin是有益的。在有氧和缺氧条件下,SGH10 可杀死大肠杆菌,但不会影响肺炎克雷伯菌的生长(图 6A)。GIE492和ICEKp10的敲除恢复了大肠杆菌的存活(图6A)。当与SGH10ΔmceA或SGH10ΔclbP竞争时,相对于SGH10,更多的大肠杆菌存活下来,这表明mccE492和colibactin通过独立的途径杀死大肠杆菌(图6A)。此外,在有氧条件下,与SGH10ΔmceAΔclbP竞争时,比与SGH10ΔmceA或SGH10ΔclbP竞争时,更多的大肠杆菌存活(图6A)。尽管GI可能含有其他使SGH10能够杀死大肠杆菌或清除铁等营养物质的因子,但这些结果表明mccE492和colibactin协同增强了colibactin的作用。
然后我们与已知定植于人类肠道的专性厌氧菌进行 HvKp 竞争。我们发现人类肠道专性厌氧菌对colibactin的作用敏感(图6B-D)。这些人类肠道分离株对应于梭状芽孢杆菌目的分类单元,这些分类单元被产colibactin的 SGH10 定植的小鼠中被耗尽。其他重要的人类肠道厌氧菌,如艰难梭菌、青春双歧杆菌和长双歧杆菌也对 mccE492 和colibactin敏感(图 6E-G)。此外,我们还与一组克雷伯菌分离株(clb-mce- 和 clbS-mceB-)进行了 HvKp 竞争。我们观察到在有氧或缺氧条件下,大肠杆菌和克雷伯氏菌对 mccE492 和 colibactin 的敏感性存在差异(图 6A 和 7A)。一些菌株在有氧条件下对mccE492敏感,但这些菌株在缺氧条件下不被mccE492杀死。一些在有氧条件下对colibactin不敏感的菌株在厌氧生长期间对colibactin敏感。在缺氧条件下,只有肺炎克雷伯菌NUH28和NUH56对mccE492和colibactin敏感(图7A和8C)。在这些实验中,肺炎克雷伯菌的生长不受与其他细菌共培养的影响(补充图 9 和 10)。
我们的筛选显示,一些革兰氏阳性菌,如艰难梭菌、青春双歧杆菌和长双歧杆菌,对colibactin和 mccE492 都敏感(图 6E-G 和 7A)。主要的革兰氏阴性人类肠道菌群多形拟杆菌对肺炎克雷伯菌均具有抗性(图 6F 和 7A)。

6.肺炎克雷伯菌不使用 mccE492 和colibactin与密切相关的分离株竞争
为了确定肺炎克雷伯菌中的mccE492和colibactin对于与密切相关的细菌猎物的竞争是否重要,NUH56在体内竞争期间被用作猎物(图7B),因为它是易于被杀死的mce-clb-ST23谱系菌株(图 7A)。我们没有观察到 NUH56 猎物与 SGH10 或其各种突变体竞争时的定植存在统计学上的显著差异(图 7C)。我们观察到colibactin的表达有利于 SGH10 定植,尽管它不会杀死 NUH56。与第11天的SGH10ΔlacZ相比,SGH10ΔclbPΔlacZ和SGH10ΔmceAΔclbPΔlacZ的粪便携带量显着减少(图7D)。第13天,相对于SGH10,SGH10ΔmceAΔclbPΔlacZ的粪便携带量减少,但SGH10ΔclbPΔlacZ没有减少(图7D)。SGH10ΔmceAΔlacZ 以及 SGH10ΔlacZ 定植于肠道(图 7D)。第13天,与用SGH10ΔlacZ和SGH10ΔmceAΔlacZ定植的小鼠相比,用SGH10ΔclbPΔlacZ和SGH10ΔmceAΔclbPΔlacZ定植的小鼠中NUH56的粪便细菌载量低约2个对数(图7D)。这些结果支持这样一种模型,即microcin和colibactin的关键靶标不是与 HvKp 密切相关的竞争者(例如其他易感肺炎克雷伯菌),而是影响定植抗性的微生物组的其他成员。事实上,colibactin和 mccE492 似乎甚至有利于其他肺炎克雷伯菌菌株(例如本身不产生这些因子的 NUH56)的定植。

7.mccE492 和colibactin对细菌猎物的影响
MccE492 是一种成孔细菌素,具有抗革兰氏阴性菌活性。我们证明纯化的 mccE492 可以以剂量依赖性方式扰动大肠杆菌的外膜(图 8A 和 B)。此外,纯化的 mccE492 对革兰氏阳性菌青春双歧杆菌、长双歧杆菌和长双歧杆菌的存活具有剂量依赖性影响。
采用TUNEL检测来检查DNA损伤,我们发现SGH10在有氧和缺氧条件下诱导TUNEL阳性大肠杆菌显著增加(图8C和D。当 TUNEL 阳性大肠杆菌与 SGH10ΔmceA、SGH10ΔclbP 或 SGH10ΔmceAΔclbP 竞争时,未观察到 TUNEL 阳性大肠杆菌显著增加(图 8C 和 D)。使用碘化丙啶(PI)摄取测定,我们证明用 mccE492 处理可增加 BA 和 BL 的通透性(图 8F 和 G)。这些结果表明 mccE492 引起 BA 和 BL 中膜的扰动。此外,我们观察到 SGH10 诱导 BA 和 BL 中 DNA 损伤显着增加(图 8G 和 H)。BA 和 BL 中肺炎克雷伯菌诱导的 DNA 损伤均依赖于colibactin和 mccE492(图 8G 和 H)。
【总结】
总的来说,我们描述了两个 HvKp 谱系中 GIE492 和 ICEKp10 之间的关联。Colibactin和 mccE492 的组合对于 HvKp 在胃肠道中的定植是有益的。Colibactin 和 mccE492 使 HvKp 能够杀死细菌肠道共生体,从而在拥挤的内源性微生物组空间中站稳脚跟。我们的结果表明,GIE492 和 ICEKp10 在定植过程中带来的好处促成了 CG23-I 谱系的主导地位。
-
点赞 (0人)
- 收藏 (0人)

