
微生物组学研究必学知识--鉴定细菌毒力基因策略
- 牛祥娜
- 4038
- 2023-05-19 18:06:05
- 汇编
鉴定细菌毒力相关因子是了解细菌致病分子基础的关键步骤。毒力基因通常指将能增强细菌感染能力、逃避宿主免疫反应并传播的基因。这些基因可编码毒素、黏附素以及酶类等因子,毒素和黏附素是比较容易鉴定的致病因子,酶类可在养分限制的环境中增强病原体的代谢过程。然而动物模型的种类、感染途径或剂量等变量可显著改变我们对个体基因对细菌致病性影响的评价。此外,宿主的免疫状态或遗传构成能够影响宿主与病原体的相互作用,也增加了评估单个基因对细菌致病作用的难度。故此,我们在认知毒力基因的概念上,可以将其理解为任何能够直接或间接提高: ①细菌吸附和/或入侵;②复制;③组织损伤;④免疫逃避;⑤易感动物宿主传播的遗传因子。
过去几年中,通过分子技术和遗传策略来鉴定细菌的毒力决定簇,研究者利用上述方法结合基因组技术可进一步探明病原菌对不同宿主的转录反应,并可确定单个毒力基因在感染过程中的作用。本文将重点介绍鉴别毒力基因的一些方法及其研究进展。
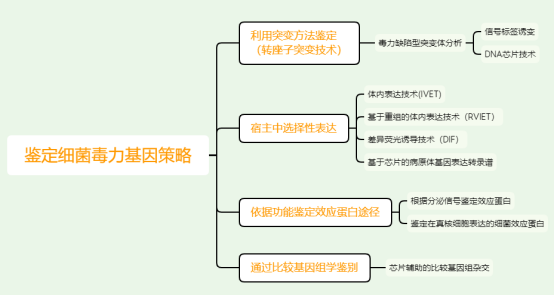
1、利用突变的方法鉴定细菌的毒力基因
转座子是目前在细菌中引入随机突变、功能缺失突变的最有效的分子工具。转座酶能够识别特定的DNA反向重复序列,同时切除这些重复序列的侧翼区,然后将催化切除的DNA片段插入到靶DNA分子的随机位点。通常插入序列会导致基因的插入性失活。可以将抗生素耐药标记或者报告基因插入到侧翼具反向重复区的序列构建出“转座子”,将上述遗传元件转运到细菌中,然后利用转座子编码的抗生素标记负选转座子插入的细菌并对其进行扩增,由于转座子可在细菌染色体的任意位点发生合,所以很容易构建出在所有非必需基因上转座子插入的细菌突变体文库。
1.1毒力缺陷型突变体的平行分析:信号标签诱变
信号标签诱变(STM)是毒力缺陷突变体平行分析的第一个系统(图)具特异性寡核苷酸条形码的转座子可追踪文库中的单个突变体。在动物体内传代后,可获得存活的突变体,扩增与转座子相关的DNA条形码,并将其作为抗合成条形码的杂交探针而固定在滤器上。通过缺失相应的条形码来评估特定突变株的缺失。
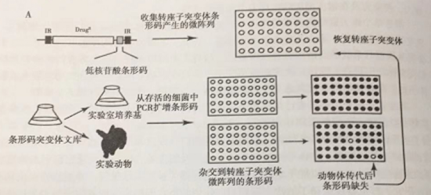
图.信号标签诱变(STM)技术
1.2 DNA芯片技术在转座突变体中的应用
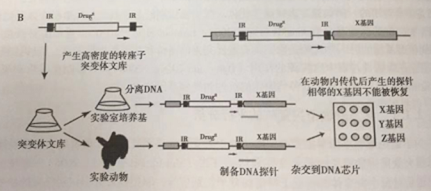
由于微生物全基因组数据的增加,将转座子插入位点相邻的DNA序列作为替代人工条形码用于追踪转座子插入位点。已经建立了一系列用于鉴定转座子插入位点的方法,包括转座子插入位点杂交技术(TRASH),转座子的微阵列追踪(MATT) ;这些方法以转座子的特异序列和随机寡核苷酸作为引物,通过PCR选择性扩增转座子插人位点周围的特有序列。首次在结核分枝杆菌中应用了这种方法,利用突变文库中的诱变菌分别感染小鼠,通过PCR扩增幸存细菌中插入位点周围的序列,并用荧光基团标记DNA探针,将输人库中突变体插位点周围序列扩增后用不同的荧光标记后作为对照组,然后将两种探针混合后与含有病原体全基因组的芯片进行杂交。就信号标签诱变来说,通过动物传代后毒力基因转座子插入的探针代表性较低。由于所有基因在芯片上的位置是已知的,那么在单个实验中就能鉴定出对细菌在动物体内生存所必需的所有基因。
2、通过在宿主中选择性表达鉴定细菌的毒力基因
细菌在感染动物体内定植、存活、复制所需的基因在宿主体内的适当时间优先表达。温度升高,降低机体铁代谢水平或存在抗菌肽等一些简单的刺激可提醒细菌已经进人宿主体内。基于这些观察,通过筛选上述条件下可以表达的基因发现了一些重要的毒力因子。然而,基因的表达是对多信号事件的响应并激活许多信号分子的结果,因此模拟宿主体内的环境十分困难。此外,宿主内的环境既不是均匀的也不是一成不变,动物体内具有不同的生态位,以及时常变化的免疫状态(例如:炎症),因此实验室很难提供这些精确的刺激范围。
在复杂的环境中鉴别细菌表达的基因可采用基因选择策略,该策略通过构建含有启动子元件及报告基因的融合载体,以利于在宿主中对细菌克隆进行选择,然后根据报告基因活性缺失在固体琼脂平板上对细菌进行体外筛选,可在宿主内激活而不能在体外激活启动子元件调控的基因被称为体内诱导基因。在过去数十年中,荧光蛋白和其他遗传标记等不同组合的选择标记已用于鉴定可在哺乳动物细胞内和动物器官内表达的病原基因。
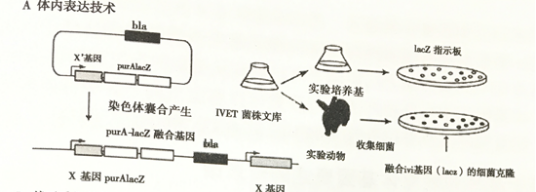
2.1体内表达技术(IVET)
为了鉴定进入动物机体后病原菌所表达的一系列基因, Mahan等人研究出了一种称为“体内表达技术”的启动子诱导技术(IVET)。构建与
purA-lacZ
报告基因融合随机启动子的菌株。
purA
基因表达是对细菌在动物体内存活所必需的,将这些通过体内表达技术产生的菌株在动物体内传代,筛选具活性启动子融合的细菌。然后在琼脂板上缩选Lacz阳性菌。大部分获得的菌株都是Lacz阳性菌,表明启动子与表达型启动子融合。少数具启动子融合Lacz阴性菌能在动物体内表达,而不能在琼脂板上表达。这些被称为体内诱导型启动子。
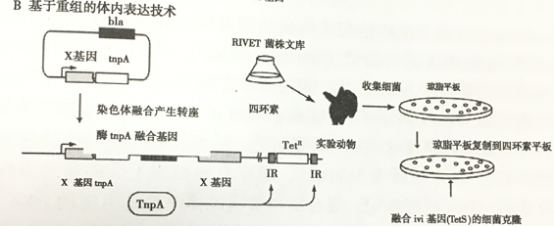
2.2基于重组的体内表达技术(RIVET)
最初的体内表达技术具有一个固有缺陷:报告基因purA必须持续在小鼠体内保持活性,因此,最初的体内表达技术在筛选过程中丢失了许多只有在特定时间或感染动物体内特定部位才表达的基因。体内表达技术策略中,报告基因对细菌在动物体内生长必需,因此扩展了这种方法的实用性。在基于重组的体内表达技术中(RIVET),转座酶(Tn—pA)作为活性基因的报告基因。在染色体的任意部位,转座酶TnpA可以识别四环素抗性基因两侧的反向重复序列。RIVET株在四环素抗性环境中可以筛选在宿主外沉默的启动子。通过动物传代后,任何活性启动子甚至是瞬时活性启动子都会导致转座酶TpA的表达并删除四环索抗性盒。从动物体筛选缺失四环素抗性的细菌。
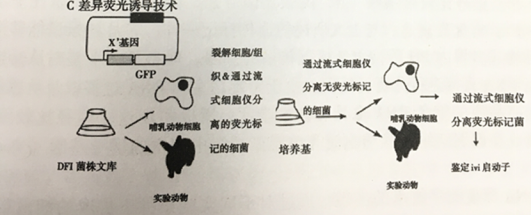
2.3荧光报告基因:差异荧光诱导技术(DIF)
绿色荧光蛋白(GFP)为一种多用途的基因表达报告蛋白,绿色荧光蛋白也能够运用于类似IVET体内表达技术的应用。通过流式细胞仪将表达不同水平绿色荧光蛋白的细菌分离开来。与 gfp融合不同强度启动子的细菌在细胞或动物上传代,然后通过流式细胞仪将荧光强度最高的细菌分离出来。然后将这些细菌暴露于实验室媒介,荧光强度最弱的细菌通过流式细胞仪分离出来。最后这些细菌通过细胞或动物传代,通过流式细胞仪再次分离荧光强度高的细菌,最后收集的细菌具有可在巨噬细胞中激活的融合启动子。通过流式细胞仪分离不同表达能力的启动子可扩展到从感染的组织直接将细菌分类。例如,从受感染的动物组织中对分枝杆菌进行分类可以鉴定出肉芽肿中特异表达的基因,肉芽肿是分枝杆菌复制的活跃部位。
2.4基于芯片的病原体基因表达的转录谱
通过检测mRNA的含量直接评估病原体表达的基因。传统的评估方法是通过基因特异性的DNA探针与总RNA进行杂交(Northerm杂交),或者用逆转录酶对总RNA处理后用特异性的基因引物进行PCR扩增。现在这种方法越来越多地应用于通过与DNA芯片杂交来监测细菌的mRNA转录本。
3、依据功能鉴定效应蛋白的途径
病原菌都存在一个特定的分泌系统,细菌的这些分泌系统能够直接将细菌蛋白运送到宿主细胞膜和细胞质中,这些分泌的“效应”蛋白负责调节许多宿主细胞的功能。通常,编码效应物的基因常与编码分泌装置成分的基因有关,或存在于病原的毒力岛上。另外,根据细菌效应蛋白与调节细胞功能的真核生物蛋白同源也可进行鉴定。然而,目前仍然有大量的效应蛋白没有被鉴定出来,目前研究集中在根据蛋白的功能特性鉴定效应物。
3.1根据分泌信号鉴定效应蛋白
细菌III型分泌系统(T3)和IV型分泌系统(T4)的靶位分别在其氨基端和羧基端,包含小的功能域。该功能域可以直接将III型(T3)分泌物和IV型(T4)分泌物易位至哺乳动物细胞中,因此,可利用融合到未知功能病原体蛋白的报告蛋白来测定这些未知功能的蛋白是否是该分泌系统的转运底物。以“信号序列”诱导方法鉴别包含T3或者T4分泌信号的蛋白,这种方法的概念与启动子诱导类似。将随机表达蛋白的细菌文库与筛选标记、报告蛋白(如β—内酰胶酶)等融合,通过监测细菌T3或T4分泌系统突变体分泌蛋白的活性缺陷以确定它们是该系统的潜在靶位。

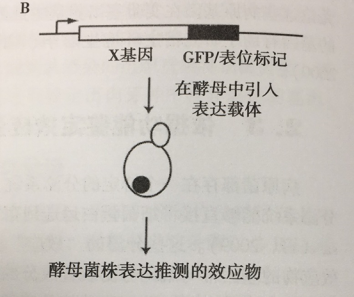
3.2鉴定在真核细胞中表达的细菌效应蛋白
通常情况下,细菌的效应蛋白具有特定的活性,可修饰或中断宿主的细胞通路。因此,细菌的这效应些蛋白在真核细胞异位表达,随后细胞产生特定的表型特征,可以提供有关它们功能的重要线索。例如,哺乳动物细胞在原生质膜、核内体、细胞核等不同亚细胞结构表达细菌效应蛋白的趋向,或者总的形态学改变和效应蛋白的细胞毒性可以帮助我们了解真核细胞中效应蛋白的靶位,这些研究工作也可作为筛选鉴定效应物的基础,由于调节膜转运、细胞骨架的动力学和信号的机制在真核细胞中是相对保守的,因此也是研究细菌病原的共同目标,酵母真菌模型能够用于筛选干扰真核细胞过程活动的细菌蛋白(Siggers and Lesser, 2008)。事实上,全细胞表型试验可检测酵母中几乎所有的信号,全细胞表型试验可筛选识别相关的分子,特别是参与宿主与病原体相互作用的分子。例如,表型实验检测细胞骨架的功能,或者在酵母的亚细胞结构适用于鉴别军团菌的效应物,这些效应物可破坏囊泡运输。
4、通过比较基因组学鉴别毒力因子
在比较基因组学中,通过在全基因组水平上比较DNA序列推断其生物学功能。例如,通过将病原菌的全基因组序列与相近的非致病性病原菌或者不同致病机制病原菌相比较,可以缩小负责致病特征基因的范围。同样,启动子序列的改变能够用于识别基因线路经过重新连接以适应宿主等新环境。
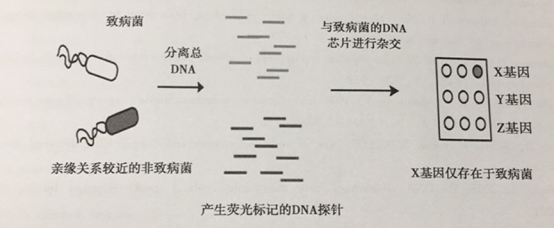
共生肠道细菌和致病性肠道细菌之间的比较基因组学揭示了在毒力菌株中(毒力岛)存在大量的毒力基因簇,这些毒力基因簇对基因水平转移事件和增强病原菌的毒力是必需的。通过芯片辅助的比较基因组杂交(CCH)对这些基因组进行大规模的比较,从可疑细菌菌株中提取的总DNA用荧光染料进行标记,并且与由菌株测序组成的有序DNA芯片杂交。对酵母的信号转导和细胞生物学通过全细胞表型分析可以鉴定筛选参与宿主与病原体相互作用的相关因子。
DNA芯片技术可比较亲缘关系较近的细菌基因组。DNA探针来源于致病菌或亲缘关系较近的非致病性菌种中分离的总DNA,用不同的荧光基团对DNA探针进行标记,然后将其与致病株的DNA芯片进行杂交,非致病性菌株缺乏的基因可能是细菌潜在的毒力决定簇。
结论
细菌的复杂性状是多个基因位点相互作用的结果,一个特定等位基因的功能主要受其遗传背景的影响。因此,将一个病原菌的毒力特性仅仅归因于一个基因的缺失或存在是不足的,有可能在致病菌和非致病菌中存在许多相同的基因作为调节细菌毒力的等位基因的组合。同样,由启动子基因导致遗传信号通路的变化可以改变暴露于宿主环境的细菌转录谱从而使得细菌病原适应共生环境。故此用于鉴别细菌毒力基因的各种分子技术、遗传学及基因组方法,在具有优势的同时也有其固有的局限性。如遗传操作适合用于病原体的常规遗传分析,而分子技术和比较基因组学方法在研究难以进行遗传操作的病原体方面非常有效。鉴于不同研究方法只能鉴别细菌的特定毒力因子,因此建议将这些技术联合起来使用。最后一点要说明的是,毒力测定方法是毒力因子鉴定技术的关键,能够忠实再现细菌入侵人体宿主细胞环境的细胞模型及动物模型,对于鉴定致病决定簇和病原学研究是至关重要的。
参考文献
【1】《Bacterial Pathogenesis·Molecular and Celluar Mechanisms》,Camille Locht.
【2】《细菌致病机制·分子与细胞水平研究》,刘永生译.
-
点赞 (0人)
- 收藏 (0人)

